Глава восьмая. Об онкогенных вирусах, природе рака и многом другом
"Бросьте все и займитесь этим!"
Сотрудники Л. А. Зильбера знали привычку своего шефа повторять эту фразу, когда какая-либо идея властно и неотступно захватывала его. Он появлялся в лаборатории в отличном настроении, энергичный, решительный, целеустремленный. В такие часы он выглядел закоренелым оптимистом, который никогда не знал сомнений, поражений и неудач.
"Бросьте все и займитесь этим!"
В данном случае "это" было некогда сумасшедшей идеей: "рак вызывается вирусами". Старая идея Л. А. Зильбера, высказанная еще в середине 30-х годов.
В 1944 году он вновь публично вернулся к ней. В докладе, сделанном на конференции Центрального онкологического института и осторожно озаглавленном "О природе злокачественных опухолей", Зильбер говорил об агенте, вызывающем злокачественные опухоли: "Какова же природа агента? Естественно было думать, что он является ультравирусом, подобным другим опухолевым вирусам типа вируса саркомы Рауса, папилломы кроликов Шоупа и других... Ряд данных и прежде всего фильтруемость действительно позволяли отнести этот агент к ультравирусам. Подобная концепция представляется мне весьма вероятной..."
Зильбер сказал все, что хотел, и хотя форма его доклада отличалась тщательно обдуманной академичностью, давние противники вирусной теории происхождения рака, стойкие оппоненты докладчика, расходились, раздраженно пожимая плечами: "Опять вирусы рака... Но кто их видел у человека?"
В самом деле, кто?
То, что вирусы могут вызывать опухоли у мышей, у крыс, у кур - доказано. Но при чем тут человек?
Загадка происхождения рака волновала ученых с тех пор, как медики научились распознавать это страшное заболевание. Что вызывает злокачественное перерождение клеток, стремительный лавинообразный их рост, когда, вторгаясь в здоровые ткани и органы, они душат, опутывают, убивают все живое?
В начале нашего века выяснилось, что рак может возникнуть под влиянием разных химических веществ, их стали называть канцерогенами. Ртуть и мышьяк, дым сигареты и анилиновые красители, каменноугольная смола и минеральные масла, типографская краска и асбест... Как писал Б. Глезмер в книге "Человек против рака" еще в 60-е годы: "Если верить некоторым авторитетам, мы живем почти исключительно в окружении канцерогенов. Подчас кажется, что правильнее всего было бы сказать: лишь горстке из нас каким-то чудом удается выжить в этом канцерогенном океане".
Итак, ливень, поток, потоп канцерогенов. А может быть, и генетический рок: есть люди, которым предопределено заболеть раком, это заложено в них наследственно, генетически?
Или причина всего в каком-то неизвестном "раковом микробе"?
Ничего удивительного в последнем вопросе нет. Он неизбежно вставал перед исследователями.
Ушел в историю XIX век с его блестящими открытиями в микробиологии. Человек нашел возбудителей многих страшных болезней - мы уже говорили об этом. В конце его Д. И. Ивановский открыл мир вирусов, а в самом начале XX века один из первых вирусологов Европы А. Боррель впервые в печати высказал гипотезу: а не фильтрующиеся ли вирусы вызывают злокачественные опухоли?
Вскоре В. Эллерман и О. Банг сообщают, что лейкозы у кур действительно могут иметь вирусное происхождение. И готовы экспериментально подтвердить это... Впрочем, тогда лейкозы не причисляли еще к злокачественным новообразованиям, так что вопрос вроде бы совершенно неясен. Неясен для всех, кроме... И. И. Мечникова. В 1910 году этот великий провидец науки печатает в газете "Русское слово" статью, в которой пишет буквально следующее: "Одна причина рака, безусловно, находится в самом организме, но другая попадает в него в виде экзогенного начала, скорее всего - вируса".
Проходит всего только один год, и ветеринарный врач П. Раус представляет доказательства вирусной природы плотной (иначе, солидной) опухоли кур, так называемой саркомы Рауса. Это открытие было сделано в 1911 году, а Нобелевская премия за него была присуждена Раусу через... полвека. Счастье, что он успел дожить до своего триумфа!
Между открытием и его признанием (и использованием) весьма часто лежат "дистанции огромного размера". Вспомним, что законы Г. Менделя были совершенно не оценены современниками и по существу переоткрыты заново через 50 лет, когда их творца уже не было в живых. Вирус полиомиелита открыл К. Ландштейнер в Вене в 1909 году, а эффективная вакцина против этого страшного заболевания появилась на свет только в 50-е годы. Вирус гриппа впервые выделен от человека К. Эндрюсом в 1930 году, эффективных гриппозных вакцин нет до сих пор, а уж Нобелевской премией за решение проблемы гриппа, как говорится, и не пахнет. Так что Раус - счастливый человек!
Впрочем, вернемся в начало века. Мелькнувшая в те годы в работах А. Борреля, И. И. Мечникова, В. Эллермана, О. Банга, П. Рауса мысль о вирусной природе рака, мысль, в значительной мере подсказанная охотой за вирусами, погасла на целую четверть века.
В самом деле, были открыты болезнетворные вирусы, вызывающие оспу, корь, грипп, свинку, желтую лихорадку, но где он, вирус рака человека, и если он есть, почему его никто и никогда не смог настигнуть и увидеть?
...14 декабря 1935 года. I Всесоюзное совещание по изучению ультрамикробов и фильтрующихся вирусов.
На трибуне - Зильбер. Советская вирусология только рождается. И он один из самых страстных ее пионеров, создатель первой специализированной вирусологической лаборатории Наркомздрава РСФСР. Ученый начинает с общепризнанных аргументов:
"Существует мнение, что фильтрующиеся вирусы редко поражают человека. Это, однако, совершенно неверно. Если мы подсчитаем заболеваемость... то получим, что... с 1929 по 1934 год... гриппом, корью, полиомиелитом и оспой заболели 25 142 650 человек, в то время как основными бактерийными инфекциями - 4 072 446 человек".
Итак, убеждать в необходимости изучения вирусов, кажется, не приходится.
Но... И далее Зильбер заговорил о том, что не могло не вызвать глубокой и настороженной тишины зала:
"...Необходимо остановиться на успехах в области изучения этиологии некоторых злокачественных опухолей. Я прекрасно понимаю необходимость чрезвычайной сдержанности и глубокой осторожности в этом сложном и важном вопросе. Однако невозможно обойти молчанием работы самого последнего времени, в которых высказывается принципиально новый взгляд на эту сложную проблему... Позволительно думать, что фактор, вызывающий некоторые опухоли млекопитающих, является не самой клеткой этой опухоли, а экзогенным, автономным от нее агентом, который в иных случаях, однако, так тесно связан с ней, что не может быть отделен от нее фильтрованием... Пройти мимо этих фактов не представляется возможным..."
Настороженность зала сменилась недоумением, недоумение разрешилось непониманием. Хорошо еще, если вежливым, без усмешек и иронических восклицаний... Слишком многим, в отличие от Зильбера, эти факты показались не стоящими внимания, мимо них вполне можно было пройти... Да и какие факты? Опухоли мышей, крыс и кроликов, вызванные искусственно в эксперименте?
И все же исторические слова тоже были произнесены: "...фактор, вызывающий некоторые опухоли млекопитающих, является... агентом, автономным от клетки, который... однако, так тесно связан с ней, что не может быть отделен от нее фильтрованием". Слово вирус еще отсутствует, но оно и только оно должно стоять здесь, рядом с другими: "не может быть отделен от нее (клетки) фильтрованием". Пока это больше намек, чем вывод, но какой вещий намек!
Моцарт говаривал о "мгновении, когда сразу слышишь всю еще не написанную симфонию". Никто не верит в нее, кроме самого автора, ибо только авторский слух готов воспринять звучащее будущее. Для этого необходим особый провидческий слух. Зильбер обладал таким слухом. И шумное неодобрение оппонентов не могло его притупить.
"Нелегко, - пишет ученик и последователь Зильбера член-корреспондент АН СССР профессор Г. И. Абелев, - постоянно быть в оппозиции к общепринятым мнениям. То, что для него было ясным и даже очевидным... отнюдь не представлялось таковым большинству исследователей... Факты, приводимые Львом Александровичем, как веские доводы в пользу своей точки зрения, далеко не всегда звучали для них однозначно... И дело здесь не в равнодушии или консерватизме. Дело в разном складе ума и в разных подходах к проблеме".
...1935 год... Может быть, слишком преждевременны идеи Зильбера? Но ведь все, что касается вирусов - возбудителей острых инфекций, - слушается со вниманием и острым интересом. И лишь соображения о фильтрующемся агенте опухолей, который "не может быть отделен от клетки фильтрованием", падают на каменистую почву... Значит, рано?
Сумасшедшая идея о вирусной природе рака или, по крайней мере, некоторых его видов подкрепляется и догадками других ученых. Н. Ф. Гамалея еще раньше высказывал идеи о том, что могут существовать вирусы, размножающиеся в клеточных ядрах. Есть ведь паразиты, которые размножаются в ядрах туфелек-инфузорий. При этом туфельки гибнут...
Аналогия еще не доказательство, размышлял Зильбер. Но опыты в поисках агента, не отделяющегося от клетки фильтрованием, увлекают его целиком, безгранично, самоотверженно.
"Бросьте все и займитесь этим!"
Опыты, сотни, тысячи опытов...
В докладе, сделанном в ноябре 1944 года на конференции Центрального онкологического института, Зильбер уже смог уверенно заявить: "Фильтраты молодых, только что возникших опухолей... оказывались способными вызвать злокачественный рост. Гипотетический экстрацеллюлярный, внесенный в клетку извне возбудитель злокачественного роста стал реальным агентом, доступным для изучения".
С 1944 года действительно было брошено все, и Зильбер со своими сотрудниками и учениками занялся только вирусной теорией происхождения опухолей. За 22 года (в 1966 году сердце Зильбера перестало биться) были проделаны десятки тысяч опытов, написаны сотни статей, сделаны десятки докладов, изданы уникальные монографии. За день до смерти ученый дописал свою последнюю, одиннадцатую (!) книгу, называвшуюся "Вирусогенетическая теория возникновения опухолей".
Но... история еще не досказана и поныне - ни история научного подвига Л. А. Зильбера, ни история раскрытия тайны рака...
Барельеф Зильбера украшает ныне стену конференц-зала института вирусологии в Москве. Здесь же профили Дженнера, Мечникова, Пастера, Ивановского...
В развитии учения о вирусном происхождении злокачественных новообразований можно выделить ряд этапов, каждый из которых характеризуется своим собственным "коэффициентом" соотношения эмпирических и теоретических компонентов.
В I период (1903 - 1910 годы) явно преобладали теоретические, точнее умозрительные, элементы, поскольку вирусный онкогенез как таковой был неизвестен, и роль вирусов в происхождении злокачественных новообразований лишь предполагалась (Боррель, Мечников).
II период (1910 - 1944 годы) характеризуется преобладанием эмпирических моментов, так как именно в это время были открыты специфические опухолеродные вирусы кур, мышей и ряда других животных.
Своеобразие III периода (1944 - 1968 годы) связано с развитием и утверждением вирусогенетической концепции Зильбера о происхождении злокачественных новообразований. Краеугольным камнем явилось положение о том, что опухолеродные вирусы представляют собой не инфекционные, а интеграционные агенты. Правомерность этой концепции была доказана сначала для ДНК-содержащих онкогенных вирусов (Л. А. Зильбер, Р. Далбекко), а затем и для РНК-содержащих онкогенных вирусов (Г. Темин). На терминах и понятиях инфекционный вирус и интеграционный вирус следует остановиться подробнее.
Инфекционный вирус-возбудитель инфекции - всегда вирулентен. Вирулентный вирус. Собственно говоря, это масло масляное. Вирус и вирулентный - слова одного корня. Вирус - яд, вирулентный - ядовитый. Вирулентный вирус - ядовитый яд. Тем не менее прилагательное вирулентный несет в себе важный и неоднозначный смысл применительно к существительному вирус.
Во-первых, этим термином определяют возбудителей острых вирусных инфекций, то есть заболеваний с коротким инкубационным периодом, бурным течением и относительно быстрым финалом: излечением, остаточными необратимыми явлениями или гибелью. Таковы грипп, полиомиелит, корь, паротит (свинка), вирусные энцефалиты, мозаичная болезнь табака и т. д. и т. п.
Собственно, рождение вирусологии как дисциплины произошло благодаря только вирулентным вирусам. Миллионные убытки заставили владельцев табачных плантаций обратиться к Д. И. Ивановскому. Тысячи искалеченных детских тел "подгоняли" вирусологов во всех странах в их охоте на вирус полиомиелита... Сейчас такая же охота ведется на возбудителя СПИДа...
Здесь сделаем важное добавление: если инфекционный вирус ослабить или убить, получится вакцина, так что инфекционный вирус может быть и не вирулентным, точнее - а вирулентным...
Вирулентный вирус, вторгаясь в клетку, подчиняет ее, заставляет работать на себя, штамповать все новые и новые копии, истощает клетку и, разграбив ее, создав полчища себе подобных, уничтожает клетку...
Так протекает острая вирусная инфекция, вызванная вирулентными вирусами.
Основной научной заслугой Зильбера является установление принципиально нового положения, согласно которому помимо вирулентных вирусов есть еще качественно отличные от них - умеренные (так называли их в самом начале, по аналогии с умеренными фагами) или интеграционные вирусы. Эти агенты вызывают образование принципиально другого комплекса вирус-клетка, в котором происходит объединение, интеграция геномов микроорганизма и клетки-хозяина.
Геном - не очень четкое понятие. Именно поэтому генетики его, как правило, не употребляют. Вирусологов оно устраивает, хотя смысл этого термина вполне генетический. Два генома в одной системе это значит: два разных хранителя наследственной информации в клетке (ДНК клетки и ДНК или РНК вируса), два самостоятельных механизма транскрипции этой информации (то есть отдельные информационные РНК клетки и вируса), два самостоятельных механизма репликации (умножения) нуклеиновых кислот - клетки и вируса, два отдельных процесса трансляции белков в рибосомах, которые делают и клеточные и вирусные белки.
Два генома и две разные судьбы. Вирус продолжается в потомстве, клетка чаще всего убита и гибнет. Геном вируса одерживает верх над геномом клетки, именно поэтому на листьях табака появляются ржавые пятна, именно поэтому оспинами изрыты лица переболевших оспой людей, именно поэтому тяжкими параличами отмечен полиомиелит: гибнут нервные клетки спинного мозга. Так бывает при взаимодействии вирулентного вируса с чувствительной клеткой. А что же происходит при интеграции геномов и что это такое вообще?
В 1935 году для Зильбера это было лишь состояние вируса, который так тесно связан с клеткой, что не может быть отделен от нее фильтрованием. Позднее речь стала идти уже не о вирусе в целом, а только о его нуклеиновой кислоте. Ясно, что это могло случиться, когда роль нуклеиновых кислот в наследственности стала наполняться современным содержанием. Вирусологи сыграли в этом важнейшем для всей биологии событии выдающуюся роль.
Еще в 1952 году американские исследователи А. Херши и М. Чейз изучали взаимодействия вируса-бактериофага T2 и кишечной палочки, помечая радиоактивной меткой либо белок, либо нуклеиновую кислоту вируса. Опыты неизменно убеждали, что в клетку кишечной палочки проникает в основном ДНК бактериофага, именно она ответственна за появление новых вирусных частиц. Не забывайте, идет только 1952 год! Уотсон и Крик еще бьются над разгадкой структуры ДНК. О роли вирусных нуклеиновых кислот в то время вообще толком ничего не знали...
И вдруг... для того, чтобы произошло заражение клетки и началась болезнь и бурное появление новых вирусов, вовсе не обязательно наличие цельного вируса, достаточно лишь его нуклеиновой кислоты. Через несколько лет этот вывод был блистательно подтвержден А. Гирером и Г. Шраммом в ФРГ и Х. Френкель-Конратом в США. Из вирусов мозаичной болезни табака удалось выделить нуклеиновую кислоту, и она одна, только она, вызывала болезнь и разрушение клеток. Открытие в 1956 - 1957 годах инфекционности некоторых вирусных нуклеиновых кислот стало весьма важной вехой в истории вирусологии.
Вскоре установили, что инфекционные процессы, вызванные нуклеиновыми кислотами и цельными вирусами, существенно отличаются, хотя эти различия носили весьма своеобразный характер. При заражении нуклеиновыми кислотами скрытый период болезни сокращался на треть, хотя объяснить - почему, никто пока не может. Предположение, что ускоренная репродукция связана с отсутствием необходимости "раздеваться", не может считаться удовлетворительным, поскольку не надо "раздеваться" только микропопуляции вируса, вызывавшей инфицирование. Но эти несколько тысяч или сот тысяч вирионов погоды не делают, их слишком мало, заболевание вызывают их последующие генерации, которые представлены уже полноценным, вполне "одетым" вирусом.
Различаются и некоторые другие реакции на цельный вирус и на голую нуклеиновую кислоту. Так, например, антитела действуют только на вирус, а вот фермент рибонуклеаза, напротив, только на вирусную РНК. Но важно подчеркнуть еще раз: различия носили непринципиальный характер.
Итак, к началу шестидесятых годов стало ясно, что молекулы РНК вируса табачной мозаики, полиомиелита, энцефалита и некоторых других обладают самостоятельной инфекционной активностью. Оказалось, что границы живого могут быть отодвинуты даже не к вирусам, а к вирусным нуклеиновым кислотам.
Но если так, неизбежно встали новые вопросы: только ли вирусная нуклеиновая кислота - единственный носитель инфекционных свойств вируса, способный осуществлять заражение без участия белка, или белок все же в естественных условиях как-то участвует в этом? Одно дело - в пробирке, in vitro, другое - в жизни.
Бурные, ожесточенные дискуссии вирусологов напоминали порой ристалища философов. В октябре 1962 года на конференции в Институте вирусологии имени Д. И. Ивановского всерьез обсуждали, что может и чего не может сделать голая вирусная нуклеиновая кислота, всегда или иногда играет свою удивительную роль и какова эта роль...
Две точки зрения столкнулись на конференции - Л. А. Зильбера и А. А. Смородинцева.
По мнению Зильбера, именно нуклеиновая кислота вирусов способствует возникновению стойких изменений наследственности клеток и разнообразных болезненных, патологических процессов. Он говорил: "Оказалось возможным, например, превратить нетоксикогенный (неядовитый) штамм дифтерийного микроба в токсигенный (ядовитый), инфицировав его фагом, выделенным из токсикогенного штамма. Нуклеиновая кислота фага, являющаяся его генетическим элементом, интегрируется (объединяется) с геномом бактерийной клетки, изменяя ее свойства и делая ее резистентной (стойкой. - Д. Г., Вл. С.) к повторному воздействию фага".
Но может ли внесение дополнительной генетической информации вызвать изменения в животных клетках? Некоторые факты заставляют думать, что круг интеграционных заболеваний более широк. Представление об опухолях как интеграционных заболеваниях создает новые аспекты изучения их патогенеза, профилактики и лечения. "Несмотря на гипотетичность некоторых суждений, - сказал Зильбер в заключение, - приведенные данные основаны на точных фактах... И это явится стимулом к широкому развертыванию исследований в новых направлениях".
А. А. Смородинцев возражал против стремления возводить вирусные нуклеиновые кислоты в ранг абсолютно самостоятельных инфекционных агентов, способных выходить из зараженных клеток и циркулировать в организме в качестве полноценных возбудителей. По его мнению, "участие нуклеиновых кислот в явлениях репродукции вирусов не дает оснований к переоценке их роли в естественном развитии острых инфекционных процессов, обусловленных участием зрелых вирусных частиц, способных полноценно проникнуть в чувствительные клетки и дифференцировать чувствительные и резистентные ткани".
Забегая вперед, скажем, что правы были оба, каждый по-своему. Через несколько лет они дали более точные формулировки, учитывающие мнения оппонентов. Зильбер подчеркнул, что острая вирусная инфекция не относится к "интеграционным болезням", а Смородинцев согласился с тем, что "роль вирусных нуклеиновых кислот может быть учтена как существенный или даже главный фактор развития вирусных опухолей..."
Это сближение крайних точек зрения объяснялось накоплением огромного количества новых фактов об особенностях разных вирусов. Оказалось, что в одних случаях нуклеиновая кислота вируса действительно индуцирует острый процесс, разрушение клеток. Это нуклеиновая кислота инфекционных (вирулентных) вирусов, в других - она интегрируется с клеточным геномом. Таковы нуклеиновые кислоты "умеренных" фагов, это было известно. Но может ли такой процесс иметь место при взаимодействии вирусов и клеток высших животных и человека?
...1964 год. Москва. Ученые, собравшиеся со всех концов планеты, отмечали столетие со дня рождения Д. И. Ивановского. Доклад Л. А. Зильбера носил странное название: "Неинфекционные вирусы". К этой необычной группе вирусов он отнес все ДНК-содержащие онкогенные вирусы. Ученый сказал тогда буквально следующее: "...можно считать доказанным, что механизм их действия на клетку заключается в основном в интеграции их нуклеиновой кислоты с геномом клетки, благодаря чему в клетке возникают наследственные изменения, выводящие клетку из соподчинения системам, регулирующим клеточный рост".
"То, что вы имеете в виду, - горячо возражал профессор В. Л. Рыжков, - есть гибридизация на молекулярном уровне. Но гибрид между человеком и вирусом немыслим, это абракадабра, с точки зрения генетика!".
Прошло несколько лет, и интеграция геномов вируса и клетки получила четкое экспериментальное подтверждение. К счастью, это произошло еще при жизни Зильбера, в середине 60-х годов, и было доказано на модели паповавирусов.
Странное название "папова" образовано из первых слогов названий ДНК-содержащих онкогенных вирусов папилломы, полиомы, вакулиолизирующего вируса (ПАПОВА).
Именно тогда было установлено, что ДНК паповавирусов действительно встраивается (интегрирует) в ДНК хромосом клеточных ядер. При этом она теряет способность самостоятельно удваиваться и давать жизнь новым вирусам. Отныне она реплицируется только вместе с хромосомой клетки-хозяина! Это и означает интеграцию двух геномов: онкогенного вируса и клетки-хозяина. Это и заподозрил Зильбер за много лет до того, как это было фактически выявлено. Именно здесь и скрыта основа основ вирусогенетической концепции происхождения злокачественных опухолей.
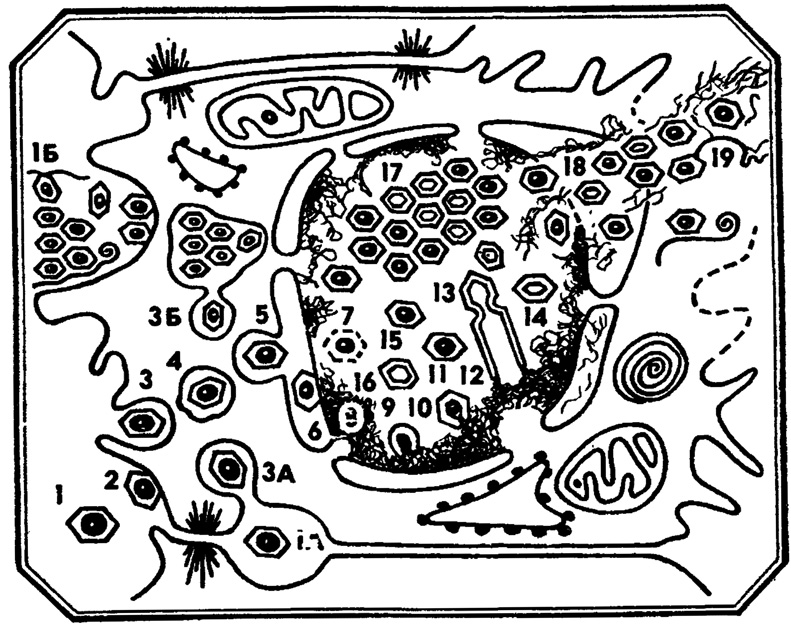
Рис. 8. Онкогенный вирус полиомы. 1 - внеклеточный вирион; 1а - внеклеточный вирион, локализующийся в межклеточном канале; 1б - внеклеточный вирион, адсорбированный на мембранных компонентах разрушенной клетки; 2 - адсорбция вириона на свободной клеточной поверхности; 3 - фагоцитоз вириона; 3а - фагоцитоз вириона из межклеточного канала; 3б - фагоцитоз вириона из гигантской внутриплазматической фагосомы; 4 - внутрицитоплазматическая фагосома, содержащая вирион; 5, 6 - проникновение вирионов в перинуклеарное пространство; 8 - внутриядерный виропласт; 9, 10 - формирование нуклеокапсидов по периферии виропласта; 11 - полный вирион; 12 - трубчатая форма вируса; 13 - предполагаемая фрагментация трубчатых форм вируса на полые 'сферические'; 14 - полные формы вируса; 15 - диффузное внутриядерное скопление вирионов; 16 - аномальная форма вируса; 17 - кристаллоподобное внутриядерное скопление полных и полых форм вируса; 18 - выход полного вирусного скопления в перинуклеарное пространство; 19 - выход вирусного скопления в цитоплазму и во внеклеточное пространство
Концепция, подкрепленная множеством фактов и опытов, стала теорией. Мы изложим самую ее суть, неизбежно кратко и упрощенно.
Итак, ДНК вируса объединилась с ДНК клетки, отныне они взаимно влияют друг на друга. Клеточная ДНК подавляет (репрессирует) ту часть вирусного генома, которая заведует синтезом структурных вирусных белков и оставляет без изменения ту часть, которая отвечает за синтез ранних белков. Поэтому производятся все новые и новые ранние белки, а структурные вирусные белки клетка вообще не продуцирует.
Роль ранних белков состоит в том, чтобы обеспечить непрерывное конвейерное производство сердцевины вируса - вирусной нуклеиновой кислоты. В драме "вирулентный вирус-клетка" ранние белки с успехом играют эту роль, обеспечивая быструю наработку многих молекул вирусной РНК (или ДНК) по "чертежу" одной проникшей молекулы.
В драме "умеренный (интеграционный) вирус-клетка" действие развертывается существенно иначе. Нуклеиновые кислоты вируса и клетки объединились, но ранние белки стараются сыграть свою обычную роль. Для удвоения (репликации) ДНК необходимо в общей сложности участие 10 разных ферментов. Но геном паповавирусов может вызвать синтез всего 6 белков. И чтобы ранние белки все же выполнили свою главную миссию, необходимо активизировать деятельность клеточных ферментов, принимающих участие в синтезе клеточной ДНК, поставив их на службу вирусам.
Но при объединении геномов вируса и клетки ферменты бездействуют, синтез структурных вирусных белков заторможен клеткой; происходит лишь синтез ранних белков, он идет беспрерывно, постоянно. При этом все время удваивается и ДНК клетки, в которой сидит ДНК вируса, что понуждает клетку к беспрерывному клеточному делению. Итак: ранние белки "подталкивают" клетку к преждевременному делению, каждое новое деление приводит к новому синтезу ранних белков, а те опять... Порочный круг замкнулся. Вирусная нуклеиновая кислота, как кнут, погоняет клеточное размножение, выходящее из-под регулирующих воздействий организма. Автономно и постоянно делящиеся клетки приобретают целый комплекс новых свойств, теряя при этом нормальный облик, становясь лично бессмертными и неся при этом гибель организму...
Повторяем, фактическое доказательство интеграции ДНК паповавирусов, прежде всего вакуолизирующего вируса (так называемого ОВ40-"О"-обезьяньего) были получены при жизни Зильбера в его лаборатории и в лаборатории выдающегося американского вирусолога Р. Далбекко. Но вопрос о том, является ли механизм интеграции обязательным и для РНК-содержащих онкогенных вирусов, оставался к 1966 году открытым.

Рис. 9. Схематическое изображение монослойных (1) и трансформированных (2) клеток
Действительно, как можно представить себе интеграцию вирусной РНК в клеточную ДНК? Молекулярная биология таких механизмов не знает. А ведь РНК-содержащие онкогенные вирусы весьма многочисленны и значительны. Достаточно напомнить, что к их числу относится знаменитый вирус куриной саркомы, об открытии которого П. Раусом в 1911 году мы говорили в самом начале этой главы.
Вирус саркомы Рауса был тщательно изучен в лаборатории Зильбера. Было установлено, что вирус этот может преодолевать видовой барьер и вызывать опухоли у млекопитающих - крыс и морских свинок. За это открытие Л. А. Зильбер и Г. Я. Свет-Молдавский, И. Н. Крюкова и А. С. Скорикова были удостоены Государственной премии СССР. Но как РНК этого онкологического "чемпиона" может интегрировать в клеточную ДНК, оставалось неясным. В вирусогенетической концепции происхождения рака зияла крупная брешь...
Правда, еще в 1964 году совсем молодой тогда ученик Р. Далбекко, Г. Темин, высказал ряд соображений о том, что геном вируса саркомы Рауса сохраняется в пораженной им клетке в форме этакого "провируса", который представляет собой не РНК, а ...ДНК! Интересная гипотеза! Но... совершенно непонятно, как РНК вируса саркомы Рауса могла превратиться (!) в ДНК. Молекулярная биология таких превращений не знала, а Г. Темин был... очень молод и крайне самонадеян.
Надо отметить, что, к счастью, с этими чертами легко сочетались поразительная трудоспособность и редчайшая целеустремленность. По сути дела, он всю жизнь занимался одним вопросом, который ему предложил еще в студенческом кружке его учитель Р. Далбекко - механизмом репродукции вируса саркомы Рауса. Первые 8 лет работы, с 1956 по 1964, привели Темина к созданию гипотезы провируса, в которую, будем справедливы, почти никто не поверил, а следующие 6 лет ушли на доказательство этой гипотезы. В 1970 году Г. Темин, работавший в Висконсинском университете, и независимо от него в Массачусетском технологическом институте Д. Балтимор обнаружили у некоторых РНК-содержащих вирусов неведомый доселе фермент, способный синтезировать ДНК-копию на матрице вирусной РНК. Статья Г. Темина, опубликованная в январском (за 1972 год) журнале "Nature", так и называлась: "Синтез ДНК, направляемый РНК". Вновь открытый фермент получил название "обратная транскриптаза" (ревертаза), а все вирусы, его содержащие, стали именоваться весьма своеобразно и даже несколько игриво - "ретровирусы". За это открытие Г. Темин и Д. Балтимор были удостоены Нобелевской премии, это было признано величайшим событием в биологии, а в вирусогенетической концепции закрылась гигантская брешь.
Мы недаром назвали эту главу столь пространно: об онкогенных вирусах, природе рака и... многом другом.
Это действительно так: начав рассказ с гипотезы Борреля о возможной роли вирусов в происхождении рака, мы буквально не можем остановиться, поток событий несет нас и помимо нашей воли заставляет касаться все новых и новых биологических проблем....
Итак, изучение первого, по-своему классического объекта онковирусологии - вируса куриной саркомы, начатое Раусом в 1911 году, привело Темина и Балтимора к открытию обратной транскрипции и нового фермента - ревертазы. Значение этого открытия оказалось столь велико, что мы вынуждены сейчас коснуться проблем, далеко выходящих за рамки и онкологии и вирусологии.
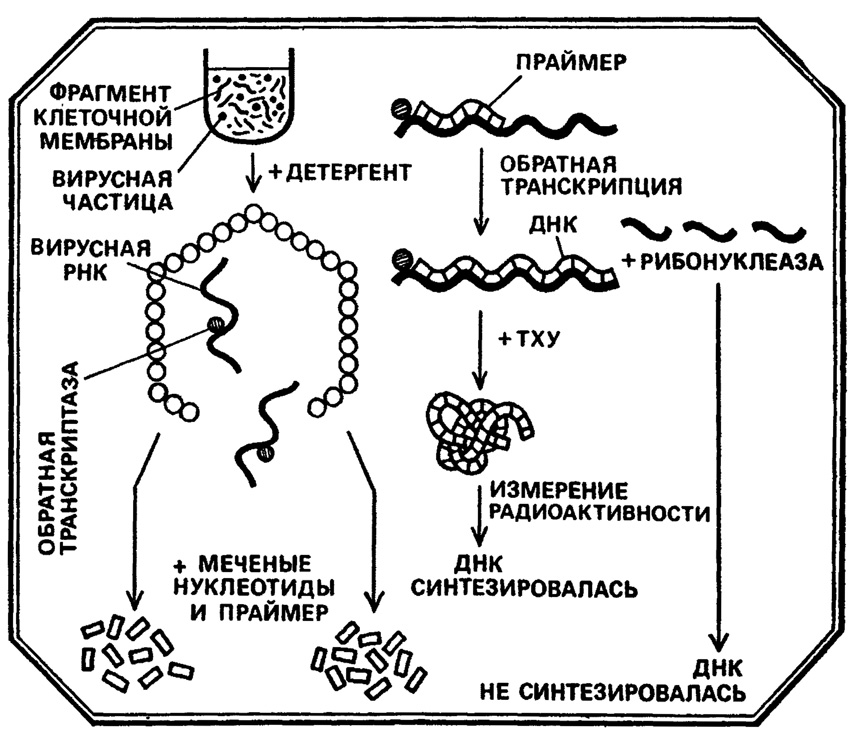
Рис. 10
Со времени выдающегося открытия Уотсона и Крика, то есть с середины 50-х годов нашего века, бесспорной считалась основная догма биологии, согласно которой гены, заключенные в двойной спирали ДНК управляют активностью любой клетки посредством двух процессов: транскрипции, в ходе которой на ДНК, как на матрице, синтезируются молекулы РНК, и последующей трансляции, в ходе которой на РНК, как на матрице, синтезируются молекулы белков. Путь ДНК→РНК→белок казался единственным, незыблемым и универсальным! РНК-содержащие вирусы казались несущественным, хотя и очевидным исключением, о котором не очень-то и задумывались... Никто, однако, не сомневался в том, что наследственная информация у этих вирусов хранится не в ДНК, а в РНК, ну а остальное происходит "почти так же", как у ДНК-содержащих вирусов. Достаточно сравнить репродукцию вируса полиомиелита и гриппа, например, чтобы в этом убедиться.
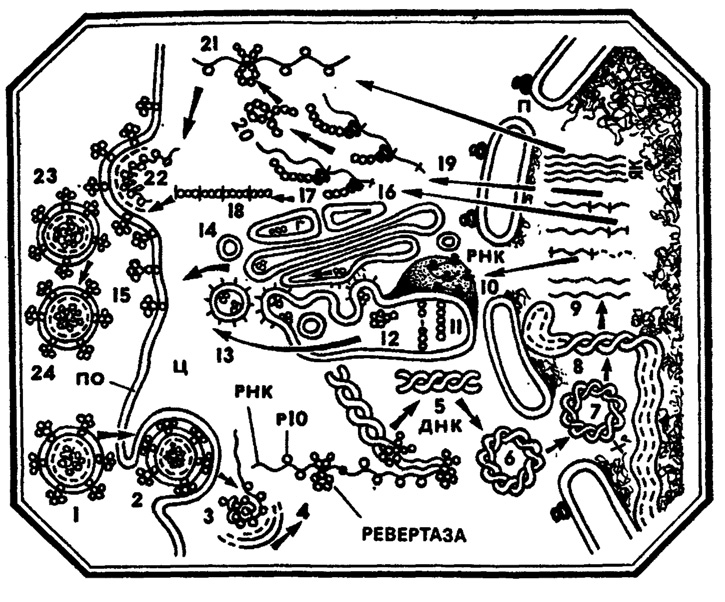
Рис. 11. Развитие онкогенного РНК-геномного вируса. 1 - внеклеточный онковирус; 2 - проникновение вируса в клетку; 3 - внутриклеточное 'раздевание' онковируса; 4 - транскриптивный комплекс; 5 - двуспиральная вирусная ДНК; 6, 7 - транспорт кольцевой ДНК онковируса в ядро клетки; 8 - интеграция ДНК-транскрипта онковируса в хромосому клетки; 9 - вирусная РНК; 10, 11 - синтез вирусных гликопротеидов на мембранах эндоплазматического ретикулума; 12, 13 - предполагаемый транспорт вирусных гликопротеидов к поверхности клетки в опушенных везикулах; 14 - предполагаемый транспорт вирусных гликопротеидов через аппарат Гольджи; 15 - локализация гликопротеидов на поверхности клетки; 16 - 18 - синтез белков сердцевины онковируса на свободных полирибосомах; 19 - 20 - синтез обратной транскриптазы онковируса на свободных полирибосомах; 21 - транспорт вирусного РНК к поверхности клетки; 22 - формирование онковируса в процессе почкования на поверхности клетки; 23 - внеклеточный онковирус типа А (С); 24 - внеклеточный онковирус типа С; Ц - цитоплазма, П - поры ядерной оболочки, ЯК - ядро клетки, ПО - плазматическая оболочка, Хр - хромосома клетки
В течение первого десятилетия после открытия Теминым и Балтимором обратной транскрипции, то есть до начала 80-х годов, казалось, что этот необычный феномен, укладывающийся в совсем иную формулу: РНК-ДНК-белок, касается только одного из семейств РНК-содержащих вирусов - ретровирусов, где он и был обнаружен, и более никого. Хотя слов нет: семейство это весьма обширно, а представители его заслуживают самого пристального внимания.
В семейство ретровирусов входит три так называевые подсемейства:
1) опухолевые вирусы,
2) вирусы - возбудители некоторых медленных инфекций,
3) так называемые "пенящие" вирусы.
К первому из этих подсемейств относятся возбудители лейкозов и некоторых плотных опухолей млекопитающих, птиц и рептилий, единственный бесспорный (пока!) возбудитель опухолевого процесса у человека (вирус так называемого Т-клеточного лейкоза людей) и агент, вызывающий... СПИД, - ни больше ни меньше! Заметим, однако, что вирус СПИДа обратную транскриптазу содержит, но опухолеродным не является: в отличие от своего близкого собрата - вируса Т-клеточного лейкоза человека - он не трансформирует человеческие лимфоциты, а убивает их.
Ко второму подсемейству относятся возбудители некоторых так называемых медленных инфекций. Мы уже упоминали об их существовании. Само понятие такого рода ввел в науку исландский исследователь В. Сигурдссон в 1954 году, высказав предположение о причине медленной инфекции - скрепи (почесухе овец).
К подсемейству пенящих относятся вирусы, не обладающие онкогенностью, но поражающие многих млекопитающих, включая человека и вызывающие так называемые синцитиеобразующие эффекты, то есть слияние клеток.
Уже из этого краткого перечня вытекает, что обратная транскрипция вовсе не равнозначна злокачественной трансформации. Для всех перечисленных агентов характерно образование ДНК-копии вирусной ДНК и встраивание этой копии в ДНК клетки-хозяина. Именно это необходимо для образования новых вирусных частиц, таков механизм репродукции этих вирусов, но последствия для клетки могут быть самыми различными: полная гибель (как при СПИДе), слияние разных клеток в многоядерный синцитий (при репродукции пенящих вирусов), и наконец - злокачественная трансформация (как при лейкозах и образовании некоторых плотных опухолей у млекопитающих, птиц, рептилий и Т-клеточном лейкозе человека).
Итак, открытая на модели онкогенных РНК-содержащих вирусов обратная транскриптаза оказалась инструментом репродукции целого ряда неонкогенных ретровирусов. И все же это была пока только вирусология, причем вирусология одного только семейства РНК-содержащих вирусов. Однако в последние годы стали появляться данные о том, что обратная транскрипция наблюдается не только при репликации вирусов, но и в незараженных клетках дрожжей, насекомых и млекопитающих.
Как это происходит и что это означает? Речь идет о целом направлении исследований в современной молекулярной генетике, и здесь очень трудно выделить самое существенное, тем более что исследования продолжаются. Но мы попробуем.
Прежде всего необходимо подчеркнуть, что в последние годы были открыты совершенно неизвестные ранее генетические элементы, присутствующие в хромосомах большинства, а по-видимому, вообще всех организмов (во всяком случае, они уже обнаружены у бактерий, дрожжей, растений, насекомых и позвоночных) - так называемые транспозоны. Эти элементы способны изменять свое местоположение в геноме клетки.
По современным представлениям, в связи с открытием транспозонов, геном представляется качественно иным, чем это было не только в эпохи Г. Менделя и Т. Моргана, но и во время открытия двойной спирали ДНК Д. Уотсоном и Ф. Криком. Геном - это не застывший кусочек некоего неизвестного вещества или даже вполне конкретной ДНК, а динамическая структура, чем-то напоминающая атом, имеющий, как известно, не только стабильное ядро, но и целый рой перемещающихся элементов (в частности, электронов) по орбитам вокруг. Именно поэтому академик Г. П. Георгиев справедливо отметил, что "ген постоянен в своем непостоянстве"!
Нет сомнений в том, что и сами транспозоны, и характер их перемещения отражают какие-то новые, неизвестные ранее стороны генетической регуляции, имеющие самое непосредственное отношение к непознанным еще законам изменчивости и эволюции живой материи. Открыта, по существу, новая глава в учении о наследственности и изменчивости, причем мы еще читаем только предисловие к ней...
Для того чтобы перемещаться, транспозоны должны иметь определенную структуру в своих концевых участках. Только таким путем они могут встраиваться в определенные участки хромосомы. Каково же было удивление ученых, когда выяснилось, что структура этих концевых участков практически любых изученных транспозонов оказалась аналогичной таковой в... ретровирусной ДНК. Сейчас это уже непреложная истина, известная в деталях, которые мы попытаемся популярно изложить.
Синтез ДНК-копии ретровирусной ДНК происходит в две фазы. На первой - обратная транскриптаза синтезирует так называемую "минус"-нить ДНК непосредственно на матрице вирионной РНК. Затравкой синтеза является при этом клеточная транспортная РНК. На втором этапе синтезируется "плюс"-нить ДНК, комплементарная первой "минус"-нити. Для этого в качестве затравки нужны короткие фрагменты вирусной РНК, отрезаемые ферментом от более длинных ее кусков.
Эксперименты, проведенные Д. Бишопом и X. Вармусом в Калифорнийском университете, а также Д. Тейлором в Институте онкологических исследований в Филадельфии, показали, что у концов ретровирусной ДНК- копии, находящейся в линейной двунитевой форме, имеются одинаковые последовательности длиной в несколько сотен пар нуклеотидов. Эти участки назвали длинными концевыми повторами - LTR (от английского long terminal region). Они оказались на обоих концах ретровирусной ДНК (а мы помним, что это и есть провирус Темина!) абсолютно идентичными с точностью до одного нуклеотида. Кроме того, у границ каждого LTR последовательности оказались также идентичными, если... если читать в противоположных направлениях. Они были названы инвертированными повторами. И наконец, было установлено, что в интегрированной вирусной ДНК по краям длинных концевых повторов располагаются одинаковые короткие последовательности, принадлежащие геному хозяина. Это означает, что при встраивании вирусной ДНК в хромосому клетки хозяина в месте интеграции происходит дупликация.
Вот каковы реальные механизмы встройки генома, РНК-содержащего онкогенного вируса в геном клетки- хозяина, процесса, предсказанного Л. А. Зильбером применительно к вирусному канцерогенезу много десятилетий назад.
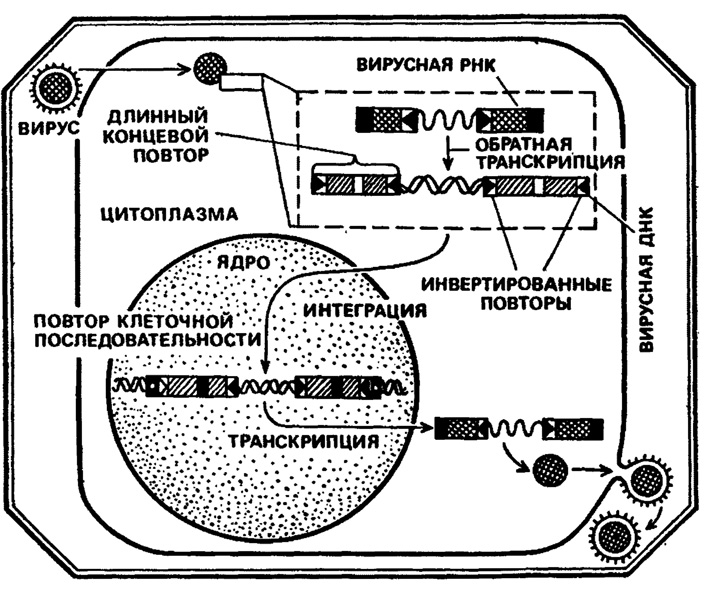
Рис. 12
Но, как мы видим, дело не только в вирусном канцерогенезе. Дело в том, что транепозоны самых разных живых существ - от бактерий до человека - также содержат LTR, инвертированные повторы и короткие дупликации последовательностей в месте интеграции. Это сходство оказалось настолько существенным, что возникла мысль о том, что обратная транскрипция может быть частью механизма перемещения некоторых транспозонов, никакого отношения, казалось бы, к ретровирусам не имеющих. В частности, особого внимания удостоился транспозон, получивший название copia и представляющий собой неинтегрированную форму ДНК в культуре клеток плодовой мушки дрозофилы, сослужившей генетике не меньшую службу, чем вирус саркомы Рауса онкологии, а вирус мозаики табака - вирусологии.
Сначала (1981 год) Э. Флавелл и Д. Иш-Горовец в Лондоне разработали метод обнаружения неинтегрированных форм ДНК copia и установили, что эта ДНК очень похожа на неинтегрированную ретровирусную ДНК в зараженных клетках. Это было очень интересно, но еще не говорило о реальном происхождении такого рода элементов. Важные сведения были получены И. Р. Архиповой в Москве, в Институте молекулярной биологии АН СССР. Она и ее коллеги с исчерпывающей полнотой показали, что и возникновение этих элементов и их перемещение в геноме осуществляется в культуре клеток дрозофилы с помощью обратной транскрипции.
Другое доказательство участия обратной транскрипции в происхождении и перемещении транспозона было получено при изучении дрожжей в лаборатории Д. Динка в Массачусетском технологическом институте. Был изучен перемещающийся генетический элемент с условным названием Ту. В серии изящных экспериментов с несомненностью было установлено, что перемещающаяся ДНК Ту является копией с... РНК, подвергшейся специальной обработке, а потому как бы меченой.
Итак, ДНК генетического фактора дрожжей, никакого отношения не имеющего к ретровирусам, является копией не ДНК, а некой РНК. Ясно, что это могло быть результатом только обратной транскрипции. Неудивительно, что перемещающиеся генетические элементы стали называть ретротранспозонами.
Итак, открыт новый неведомый ранее способ синтеза генов живых существ самого различного уровня организации: от ретровирусов до млекопитающих. Таким способом является обратная транскрипция ДНК на матрице РНК. Важно подчеркнуть, что синтез ДНК- копий ретровирусной РНК является не более чем частным случаем такого рода процесса, хотя он и был открыт первым.
Это неудивительно, но знаменательно: началась же вся вирусология с открытия вируса мозаичной болезни табака - не единственного, да и не главного представителя царства Вира (особенно в наш век тотальной борьбы с курением!). Похоже, что и обратная транскрипция ретровирусной РНК также не самая главная заслуга этого ферментативного механизма.
Есть мнение (его автор - американский молекулярный биолог Дж. Дарнелл-младший), что первым значимым веществом наследственности была именно РНК, а отнюдь не ДНК. Этому взгляду способствует большая универсальность РНК: она способна хранить информацию и воспроизводиться, в сущности, так же, как ДНК, но в отличие от нее может непосредственно направлять синтез белка и даже вести себя как фермент, о чем мы уже говорили в первом разделе книги.
Все это очень интересно и весьма вероятно. Но даже если все это так, то бесспорно, что на каком-то этапе развития жизни РНК передала свою роль ДНК. Именно поэтому обратная транскрипция является древним механизмом переноса информации с РНК на ДНК. Так, процесс, который вначале рассматривали как экзотическую способность, присущую лишь небольшой группе вирусов, теперь помогает проследить путь эволюции, приведшей к жизни на основе ДНК.
Из всех этих данных вытекают важные следствия по всем обсуждаемым нами вопросам.
Во-первых, становится совершенно очевидным, что нельзя говорить о каком-то общем происхождении всех вирусов. Видимо, из-за этого уязвимы все рассмотренные выше концепции. Кажется бесспорным, что РНК-содержащие вирусы - более древние, а ДНК-содержащие - более молодые. Ведь вирусная РНК в значительной степени сохранила все признаки древней РНК, которые у нее были, когда жизнь на основе ДНК еще не существовала: РНК - и хранитель генетического кода этой группы агентов, и непосредственный организатор синтеза вирусных белков в рибосомах инфицированных клеток, а в целом ряде случаев (применительно к малым рибовирусам) и монопольный организатор всего инфекционного процесса (вспомним инфекционность РНК вирусов полиомиелита, арбовирусов и т. д.).
Можно предположить, что эти рибовирусы являются истинными потомками тех древних доклеточных форм жизни, когда ДНК еще не стала практически монопольным хранителем генетической информации почти всего живого. В рамках этой гипотезы вполне вероятным кажется представление об определенном эволюционном усложнении древних рибовирусов от пикорнавирусов через арбовирусы к парамиксовирусам и ортомиксовирусам. В этом ряду можно отметить увеличение массы рибонуклеиновой кислоты, соответственно количества вирусиндуцируемых белков, появление отдельных ферментов в структуре вириона и, наконец, сегментацию самого генома, что увеличивает возможности генетического маневрирования. Самый сложный в этом ряду - вирус гриппа с 8 фрагментами РНК и целым набором ферментов, с выраженной способностью изменять свою антигенную структуру и биологические свойства, возможно, и один из самых "молодых"...
Другая группа агентов - ДНК-содержащие вирусы очень похожи на "взбесившиеся" гены. По крайней мере Д. Б. Голубев склонен считать их таковыми.
Еще в 1970 году были опубликованы работы американца Субак-Шарпа, который сравнил состав молекул нуклеиновых кислот трех разных групп ДНК-содержащих вирусов и соответствующих тканей позвоночных, в которых эти вирусы паразитируют. Он установил, что четыре мелких вируса из группы папова (СВ-40, полиомы, папилломы Шопа и папилломы человека) имеют нуклеотидное строение и состав, во многом сходные с составом тканей человека и животных. Эта схожесть создает почву для объединения ДНК вируса и клеток и лежит в основе онкогенности этих вирусов. Особенно отчетливо выявилось это тем же Субак-Шарпом при изучении аденовирусов, среди которых есть, как известно, и высокоонкогенные и практически неонкогенные представители. Так вот, у высокоонкогенных аденовирусов гомология вирусной и клеточной ДНК выражена в высокой степени, а у неонкогенных - вообще не выражена.
Все ли ДНК-содержащие вирусы - взбесившиеся фрагменты клеточных ДНК или только малые (если это вообще так), мы не знаем. Возможно, раз возникнув, такого рода агенты потом самостоятельно прогрессивно эволюционировали от паповавирусов к вирусам группы оспы - сложным, большим, богато оснащенным ферментами (почти бактериям).
Ни один из нас не готов настаивать на том, что высказанные предположения бесспорны. Но каково бы ни было реальное происхождение всех этих агентов (как мы заметили, за исключением ретровирусов), они, по мнению Д. Б. Голубева, живые существа, имеющие свою эволюционную историю и "борющиеся" за выживание своего вида по всем законам дарвиновской биологии.
А что же ретровирусы? Чтобы сказать что-то об их происхождении, нужно сделать новый экскурс в область онковирусологии, в ее современное состояние.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://nplit.ru/ 'Библиотека юного исследователя'