Глава седьмая. Многоактная пьеса "Клетка и вирус" (Часть вторая. "Вирус")
В 1963 году в Таврическом дворце в Ленинграде проходила сессия Академии медицинских наук. В ее повестке стоял один-единственный вопрос: современное состояние вирусологии. В докладах были проанализированы наиболее важные разделы общей и частной вирусологии. Громко и значительно прозвучали новые тогда молекулярно-биологические термины, и среди них наиболее часто: "вирусная репродукция". Именно этот термин вызвал сильное раздражение одного из крупнейших эпидемиологов страны, маститого академика, внесшего неоценимый вклад в теорию и практику борьбы со многими инфекционными болезнями. "Почему репродукция? - грозно вопрошал он. - Почему нужно обозначать этим словом процесс, который все привыкли называть размножением? Не есть ли это дань новомодным устремлениям с целью сокрыть истинный смысл?"
Такую реакцию можно понять. Внешне - с точки зрения биолога и врача (в частности, и эпидемиолога), между размножением вирусов, бактерий, простейших, рыб и любых других организмов - нет никакой принципиальной разницы. Особь родит подобное себе дитя! Точность, с которой это подобие поддерживается в мире вирусов, нисколько не меньшая, чем во всех иных группах живых особей. Ведь заражение ребенка вирусом кори никогда не приводит к "ошибочным" результатам, ни один ребенок никогда при таком заражении не заболел не корью (а гриппом или свинкой, например!). Так не бывает! Особь порождает себе подобное дитя. Повторяем: внешне в мире вирусов происходит так же, как и в других царствах живой природы. Почему же этот процесс нужно обозначать другим термином?
Прежде чем ответить на этот вопрос, необходимо отметить общие механизмы, которые лежат в основе любого вида размножения - и вирусного, и невирусного. Это матричный синтез. Именно он определяет биосинтез белков и нуклеиновых кислот. Его отличительная черта в том, что сама матрица, содержащая интегративную информацию в виде химической структурной записи, не входит в состав конечного продукта, возникшего под ее руководящим воздействием. По образному выражению В. А. Энгельгардта, матрица подобна "знаменитому мавру", который "сделал свое дело и может уйти".
Матричный синтез - основа биологической интеграции. Из полнейшего термодинамического хаоса (включающего, в частности, смесь нуклеотидов и аминокислот) он обеспечивает получение заданного и строжайше соблюдаемого порядка - химической структуры молекулы соответствующего полимера.
Все это так. Но если говорить о размножении любых клеточных организмов, то нельзя не отметить следующей важной особенности: матричный синтез всегда сочетается там с материальной преемственностью материнских и дочерних элементов. Мы только что это подробно разобрали. Обратим здесь лишь внимание на самое важное: материнская клетка перед делением удваивает генетические запасы, а затем разделяет их поровну между дочерними клетками. Материальная преемственность, таким образом, налицо.
В той или другой форме эта преемственность наблюдается при всех видах размножения и одно- и многоклеточных организмов. Это же касается и полового размножения: в дочерний организм попадают материальные частицы родительских половых клеток. Таким образом, матричный синтез, хотя и лежит в основе воспроизводства всех биополимеров, в том числе и самой ДНК, но не является единственным условием размножения клеточных организмов, поскольку сочетается с материальной преемственностью материнского и дочернего тела. Это и есть размножение.
Ну а вирусы? Снова повторим: внешне - все то же самое, но если вглядеться внимательней (что и сделала молекулярная биология вирусов за последние три десятка лет!), оказывается: матричный синтез в этом случае есть, а материальная преемственность между "родителями" и "детьми" полностью отсутствует. И это вирусная репродукция. Самое главное в ней то, что вирусная нуклеиновая кислота не только несет в себе запас наследственной информации, но и является непосредственным инструментом ее выражения, выступая в роли инициатора процесса раскрытия генетического рода. Вот почему на сессии, с описания которой мы начали этот раздел, на реплику академика-эпидемиолога: "Почему репродукция, а не размножение", другой академик - молекулярный биолог ответил: "Именно репродукция, и никак иначе!"
Итак, вирус - не обычный микроб, который просто размножается. Вирус - это особый биологический индивид, этакий монстр, который вроде бы и размножается, но в то же время делает это все совсем не так, как все другие живые существа. В итоге новые поколения вирусов - по существу не дети поразивших клетки материнских, родительских особей, а безусловные их братья и сестры.
Столь своеобразная форма создания себе подобных, естественно, связана с молекулярной организацией вирусной частицы. По современным представлениям, вирусы не столь очевидно голы и безоружны, как это казалось после работ У. Стенли с кристаллами нуклеопротеида ВТМ (вируса табачной мозаики) середины 30-х годов. В составе разных вирусов найдено некоторое количество ферментов. И тем не менее вирус без клетки размножаться не может: и ферментов недостаточно, и, главное, нет рибосом, то есть "фабрик" для синтеза своего собственного белка. Отсутствие рибосом характерно, правда, еще и для риккетсий и хламидий - ближайших "соседей" вирусов по иерархической лестнице микромира. Именно поэтому все они (и вирусы, и риккетсии, и хламидии) не растут на искусственных питательных средах, как бактерии, плесени или грибки и используют для своего размножения живые полноценные клетки.
Вот почему так трудно выделить от больного человека, животного или растения живой вирус, вот почему содержать вирусологическую лабораторию намного дороже, чем бактериологическую. Ведь и куриные эмбрионы, и клеточные культуры, и лабораторные животные намного дороже искусственных питательных сред, в которых накапливаются бактерии. И не только накладнее, но и хлопотнее, хотя бы потому, что животных надо кормить регулярно, даже в выходные дни, а автоматов для этого пока не придумали. Да и помещений вирусологической лаборатории нужно значительно больше, и требования к их обустройству более жесткие.
Намного дороже содержать вирусологическую лабораторию не только из-за животных, но и оборудования. По этому показателю ее и сравнивать с бактериологической нельзя. Электронные микроскопы, ультрацентрифуги, сцинцилляционные счетчики стоят в сотни раз дороже, чем самая совершенная аппаратура для исследования бактерий.
Будет ли так всегда? Не станет ли возможным биосинтез вирусов в пробирке (in vitro) без клеток? В принципе такого можно ожидать, но это будет означать, что в пробирке удалось воспроизвести условия, которые реально существуют только в целой, да еще чувствительной к данному вирусу клетке. Дело это огромной сложности и неменьшей важности.
Именно об этом говорил на уже упоминавшейся нами сессии Академии медицинских наук СССР в 1963 году академик Л. А. Зильбер: "Только с созданием системы бесклеточного синтеза вируса отпадут опасения в том, что с вирусными вакцинами в организм человека не попадет при иммунизации нечто постороннее. Такая система будет знаменовать собой подлинную революцию в вирусологии".
За прошедшие с того времени 25 лет мы еще не дожили до такой революции. Клетка остается пока единственным монопольным "хозяином" вирусного размножения. Вне клеток животных, человека, куриных эмбрионов, тканевых культур репродуцировать вирус пока не удается, и это делает систему вирус-клетка в известном смысле слова центральной фигурой вирусологии. Неудивительно, что изучению взаимодействия вирусов и клеток посвящено огромное количество научных работ, которые в своей совокупности составляют главу в современном учении о вирусах. Даже генная инженерия принципиально не изменила этой проблемы. Она лишь создала возможность синтезировать вирусные белки в клетках микроорганизмов. Но опять-таки в клетках!
Как же происходит вирусная репродукция?
Мир вирусов огромен и многолик. Только непосвященным вирусы кажутся одинаковыми существами. На самом же деле, сказав вирус, мы почти ничего конкретно не сказали, так же, как если бы произнесли слово животное, не уточнив, какое животное имели в виду. Между вирусами полиомиелита и оспы, наверное, не меньше различий, чем между рыбами и птицами.
Но в воспроизводстве, размножении, а точнее - репродукции вирусов, и мы уже говорили об этом, есть важные общие черты. Именно это в значительной степени и делает разные вирусы членами одного биологического сообщества - царства Вира и позволяет в учебнике по вирусологии следующим образом подразделять главу, называющуюся "Репродукция вирусов":
адсорбция;
проникновение вирусов в клетку;
транскрипция;
трансляция;
репликация;
сборка вирусных частиц;
выход вирусных частиц из клетки.
Итак, взаимодействие вируса с клетками - многоэтапный процесс, и первым из них является адсорбция. Но прежде чем ее характеризовать, позволим себе немного пофилософствовать.
В известном смысле любое соприкосновение вирусной частицы с клеткой может рассматриваться как взаимодействие. Нет никаких сомнений, что такого рода взаимодействий было несметное множество, даже когда мы писали эти строки, а любой (и каждый) наш читатель удостаивал их своим вниманием. Однако и мы дожили до конца работы, и читателю ничто не помешает дойти до последней страницы нашего скромного труда, а это свидетельствует о том, что никаких пагубных биологических последствий такое взаимодействие не вызвало.
К сожалению, взаимодействие вируса и клеток не всегда заканчивается столь благополучно. Контакт вируса с клеткой может привести (и часто приводит!) к возникновению весьма своеобразного биологического монстра - системы вирус-клетка, к которой как нельзя лучше подходят . все упоминавшиеся нами рассуждения Берталанфи об "и" и его роли в интеграции.
Мы уже говорили об удивлении как основе познания, без чего невозможно ничего открыть и, по-видимому, нельзя быть ученым. Лауреат Нобелевской премии Д. Кендрью, открывший молекулярную структуру белка миоглобина, отмечает, что именно удивление было его первой реакцией на рентгенограмму человеческого волоса. И было настолько сильным, что послужило первым толчком к занятиям биологией, причем именно изучению больших молекул, входящих в состав живых организмов.
Система вирус-клетка не может не удивлять. В ней причудливо переплелись свойства двух самостоятельных организмов, это своего рода гибрид, химера, странный, но достаточно стойкий комплекс, ни в коей мере не сводимый к слагающим его компонентам.
Несмотря на огромное разнообразие клеток и вирусов, их взаимодействие друг с другом протекает по одному из следующих типов: автономному или интегративному... Мы написали эти термины и осеклись, как это было уже неоднократно... Как безжалостно сухи бывают слова, кратко обозначающие целые вехи истории. Два типа взаимоотношений вирусов и клеток: один и другой. А какими словами обозначить тот путь, который прошла наука, а точнее и проще - люди науки, прежде чем мы смогли так просто написать эту фразу...
Мы хорошо знаем о том, что честь открытия первого вируса принадлежит сотруднику кафедры физиологии растений Петербургского университета Д. И. Ивановскому. Он был командирован в Крым (случилось это в конце прошлого века), чтобы узнать, почему гибнут плантации табака. Сначала на табачных листьях проступали жилки, потом - мертвенные пятна; листья покрывались зелено-желтой мозаикой и опадали на землю...
Ивановский решил выделить микроб опустошительной табачной мозаики. Он процеживал заразный сок через фарфоровые фильтры с мельчайшими отверстиями: они пропускают жидкость, но задерживают бактерии. Бактерий не было, а процеженный сок (отфильтрованная жидкость) оставался столь же заразным. Что это - сверхмалый микроб? А может, просто ядовитая жидкость? Ивановский настаивал: это особые сверхмалые микробы, именно поэтому они проходят через фильтры.
Позднее их назовут фильтрующимися вирусами. В электронном микроскопе получат первые фотографии "дедушки царства Вира" - вируса табачной мозаики. Но это будет позднее.
А с самого начала было ясно, что новый сверхмалый микроб не может размножаться без клеток. Это было очевидно, но прошло очень много лет, пока стало ясно содержание драмы "вирус - клетка". Впрочем, хватит философствовать, наша драма многоактна. Акт первый: адсорбция.
Строго говоря, адсорбироваться вирусы могут (и безусловно, это делают!) не только на клетках, а... на чем угодно, в силу электростатического взаимодействия положительно и отрицательно заряженных группировок, расположенных на поверхности вируса, с одной стороны, и на любой поверхности, в том числе и клеточной - с другой. Ясно, что этот процесс неспецифичен. Но... вирус гриппа заражает человека, попав на слизистую верхних дыхательных путей, и совершенно безопасен при нанесении на кожу, например. Полиомиелитом нельзя заразиться иначе, чем проглотив его возбудителя и т. д. В чем тут дело?
За неспецифической стадией адсорбции наступает высокоспецифическая, когда белки поверхности вируса (так называемые прикрепительные) соединяются со специфическими группировками (рецепторами) на плазматической мембране клетки, если, конечно, такие рецепторы на этих клетках имеются. Для вируса гриппа, например, на слизистой верхних дыхательных путей они есть, а на коже нет. Именно поэтому грипп - респираторная инфекция, а полиомиелит - кишечная (во всяком случае - по методу заражения или, как говорят эпидемиологи, по входным воротам инфекции).
Мы не будем рассматривать структуру рецепторов на разных клетках и свойства прикрепительных белков представителей разных рибо- и дезоксирибовирусов. Скажем лишь: что в своей совокупности эти сведения достаточно точно и полно объясняют явление тропизма вирусов к одним тканям, устойчивость тех или иных клеток и вирусам и, наконец, видовую устойчивость или, наоборот, чувствительность к вирусным инфекциям. Почему, например, пикорнавирусы (возбудители полиомиелита и полиомиелитоподобных инфекций) поражают только приматов, а тогавирусы (возбудители клещевого и комариного энцефалитов) - и млекопитающих, и членистоногих, и человека? Да потому, что соотношение структуры прикрепительных вирусных белков и клеточных рецепторов не позволяет пикорнавирусам выйти за пределы отряда приматов, а тогавирусам дает возможность легко перешагнуть границы типов и проникать в клетки как позвоночных, так и беспозвоночных.
Итак, адсорбция прошла. Что же дальше? Дальше - проникновение вируса в клетку. Исторически сложилось представление о том, что вирусы могут проникать в клетки либо путем так называемого виропексиса, либо путем слияния вирусной и клеточной мембран. Вряд ли правильно говорить либо - либо. По-видимому, оба эти механизма не столько исключают, сколько дополняют друг друга. Понятно также, что каждая система вирус-клетка характеризуется своим индивидуальным способом проникновения нарушителя границы клеточного государства.
Современные руководства по вирусологии буквально переполнены описанием подробностей каждого такого отдельного разбоя, и только в некоторых из них содержатся отдельные попытки прописать лекарства, защищающие клетки от гибели. Мы имеем в виду химиотерапевтические средства, к сожалению, крайне немногочисленные в вирусологии. Но об этом - в другом месте. Сейчас в нескольких словах охарактеризуем самое главное - проникновение вируса в клетку.
Термин "виропексис" предложил в 1948 году видный вирусолог австралиец венгерского происхождения Ф. де Сент-Гроот. Это не что иное, как разновидность пиноцитоза (или эндоцитоза), то есть обычного механизма поступления в клетку из внешней среды питательных и регуляторных белков, гормонов и других высокомолекулярных веществ. В отдельных участках цитоплазмы, в особых ямках, где располагаются специфические рецепторы, после взаимодействия с вирусом возникают выпячивания (инвагинации). Затем эти выпячивания замыкаются и образуются внутриклеточные вакуоли, содержащие в себе проникшее в клетку вещество, в частности, вирус, если это был он.
Вакуоли сливаются между собой, образуя крупные внутриклеточные полости, соединяющиеся затем с лизосомами. Вы помните об эти стражах постоянства внутренней среды клеток, наполненных разнообразными ферментами? Теперь мы их видим в работе: содержимое лизосом изливается в вакуоли, и начинается эффективная ферментативная переработка. Белки разлагаются до аминокислот, нуклеиновые кислоты до отдельных мононуклеотидов и т. д. Казалось бы, лизосомы должны в порошок растереть непрошеных гостей, ведь вирусы - типичные нуклеопротеиды! Но этого не происходит. Напротив, лизосомы своими ферментами помогают вирусной мембране слиться с клеточной, выстилающей внутреннюю полость вакуоли. Вот почему мы говорили о том, что механизмы виропексиса и слияния мембран вируса и клетки не противоречат, а дополняют друг друга.
Итак, мембраны слиты! В одних системах вирус - клетка это известно до мельчайших подробностей, в других - только начинает изучаться. Например: какой из вирусных белков является белком слияния? По-настоящему он идентифицирован только у пара- и ортомиксо-вирусов. Так, у вируса кори или парагриппа (оба - парамиксовирусы) им является один из двух гликопротеидов, находящихся на поверхности вирусной частицы. У вируса гриппа (ортомиксовирус) функцию белка слияния выполняет малая субъединица гемагглютинина - важнейшего поверхностного антигена этого возбудителя. Так или иначе, но этот процесс происходит при проникновении в клетки любых вирусов.
Итак, все сказанное подводит нас к главнейшему событию, без которого не может начаться вирусная инфекция - "раздеванию", высвобождению вирусного генома от белкового покрытия. Условия для такого раздевания создаются всеми предшествующими, описанными нами процессами: связыванием со специфическими рецепторами, проникновением в клетку, слиянием мембран.
Не отвлекаясь на частности (а великое множество их уже известно и еще более неизвестно), можно сказать, что вирус, проходя внутрь клетки, раздевается постепенно, сбрасывая по мере своего взаимодействия с клеткой одну одежду за другой, стремясь обнажиться до такой степени, которая позволит ему начать смертельный для клетки "танец": для одних вирусов такими продуктами являются сердцевины вирионов, в других - нуклеокапсиды, в третьих - нуклеиновые кислоты. Но во всех случаях критерием достаточной для начала инфекции степени раздевания вируса является способность начать транскрипцию.
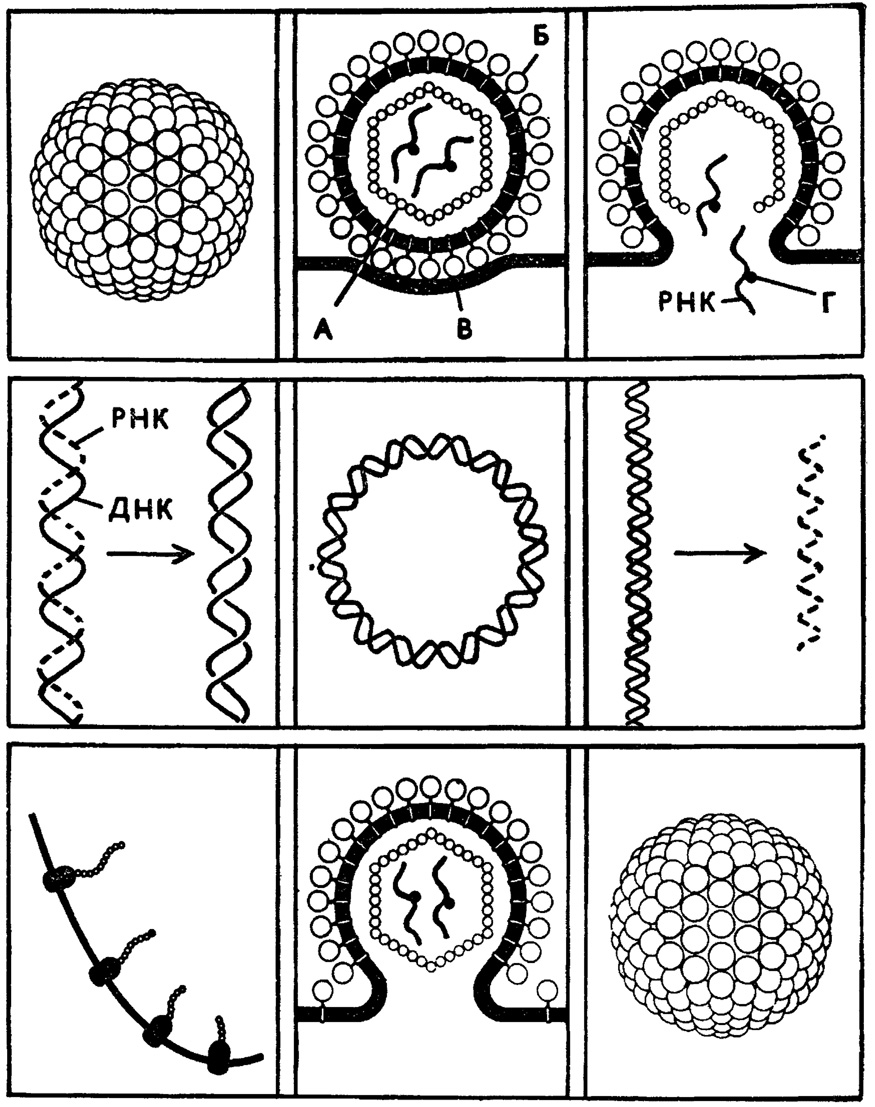
Рис. 5. Жизненный цикл ретровируса. А - белок сердцевины; Б - гликопротеин оболочки; В - клеточная мембрана; Г - обратная транскрипция
Нам не нужно здесь объяснять, что такое транскрипция - мы говорили об этом выше, путешествуя по клетке. Напомним, что центральная догма молекулярной биологии клетки выражается формулой: ДНК (транскрипция)→РНК (трансляция)→белок.
То, как реализуют свою генетическую информацию вирусы, называют - стратегией вирусного генома (!). (Заметим: это не наше выражение, мы заимствуем его из обширной литературы по молекулярной вирусологии.) Стратегия генома ДНК-содержащих вирусов в отношении синтеза иРНК та же самая, что и у клетки (ДНК→РНК→белок). Стратегия та же! А тактика?
Тактика различна у разных групп вирусных агентов. Большинство ДНК-содержащих вирусов, во всяком случае все, репродукция которых происходит в ядре (паповавирусы, аденовирусы, вирусы герпеса), используют для своей транскрипции клеточный фермент полимеразу (точнее: ДНК-зависимую РНК-полимеразу). Именно она на матрице вирусной ДНК строит комплементарную ей информационную иРНК. Вот она - молекулярная структура вирусного паразитизма, молекулярное выражение разбойного поведения вируса, проникшего в клетку-государство и использующего его "правительственный аппарат" для своих злодейских целей!
Большие ДНК-содержащие вирусы (оспы, например) размножаются не в ядре, а в цитоплазме пораженных клеток, и в силу этого не могут для своей транскрипции использовать клеточный ядерный фермент - полимеразу. Но у них есть свой собственный (вирионный) фермент ДНК-зависимая РНК-полимераза, которая и начинает транскрипцию. Заметим, что это, конечно, показатель большей самостоятельности, чем та, которая есть у "ядерных" ДНК-содержащих вирусов. Но разбойный паразитический нрав этого агента нисколько не тише, чем у его более мелких по размерам собратьев. У него, как и у них, нет своих рибосом, а потому он не может сам синтезировать свои белки и является таким же клеточнозависимым паразитом. Но на этапе транскрипции, причем именно на самой ранней ее стадии, он более автономен, имея в своей структуре некоторый "набор отмычек", если хотите.
Иначе обстоит дело с транскрипцией РНК-содержащих вирусов (рибовирусов). Здесь хранителем генетической информации является не ДНК, а другая нуклеиновая кислота, РНК. Ясно, что центральная догма молекулярной биологии ДНК→РНК→белок здесь не может применяться, как говорят физики, по определению. Как же тут происходит транскрипция? Очень по-разному, у разных групп рибовирусов. А то, что они очень разные, вытекает хотя бы из того, что по структуре и свойствам своего генома они делятся на несколько очень разных групп.
Группа первая. Рибовирусы с позитивным геномом (или плюс-нитевые вирусы). Сюда относятся пи- корна-, тога- и коронавирусы. У этих агентов сам геном, сама вирионная РНК выполняет функции иРНК. У них вообще нет транскрипции! Формула их жизни необычайно проста: РНК→белок.
У второй группы рибовирусов геном не может выполнять функций иРНК. Это так называемые минуснитевые вирусы (или вирусы с негативным геномом). Сюда относятся ортомиксовирусы (вирус гриппа), парамиксовирусы (корь, парагрипп, паротит и др.). У этих агентов транскрипция имеет место. При этом иРНК строится на матрице вирионной РНК и является, естественно, комплементарной ей. Передача генетической информации у этих вирусов происходит по формуле РНК→РНК→белок.
Фермента, который мог бы синтезировать РНК на матрице другой РНК, то есть РНК-зависимой РНК-полимеразы, в клетках нет и быть не может (мы помним, что в ядрах клеток происходит постоянный синтез иРНК, но на ДНК-вых матрицах, и делает это фермент ДНК-зависимая РНК-полимераза). Этот фермент вирусы с негативным геномом приносят с собой, оказывается, он есть в структуре вириона, это вирионная транскриптаза, или вирионная РНК-зависимая РНК-полимераза. Такие ферменты были открыты в составе парамиксо- и ортомиксовирусов в 70-е годы, и это открытие сделало понятным, как может реализоваться формула РНК→РНК→белок.
Наконец, к рибовирусам относятся и так называемые ретровирусы, в составе которых есть уж и вовсе диковинный фермент РНК-зависимая ДНК-полимераза, которая строит ДНК по матрице... РНК. Эти вирусы живут по формуле РНК→ДНК→РНК→белок.
О ретровирусах, РНК-зависимой ДНК-полимеразе (обратной транскриптазе, или ревертазе) и удивительной формуле, дополнившей основную догму молекулярной биологии Уотсона и Крика, мы будем говорить особо. Отметим только: иРНК ретровирусов гомологична геномной вирионной РНК (а не комплементарна ей, как у вируса гриппа, например), а потому ретровирусы так же, как и пикорнавирусы, относятся к плюс-нитевым. Но сколь разные структуры и свойства скрываются за этим подобием! Поистине даже подтип рибовирусовтаит в себе гигантское многообразие семейств, родов и видов по самым кардинальным молекулярно-генетическим параметрам. А ведь есть еще и второй подтип - дезоксирибовирусов: ДНК-содержащие вирусы!
Сначала отметим, что представители самых крупных по размерам вирионов ДНК-содержащих вирусов - вирусов оспы и иридовирусов - репродуцируются в клеточной цитоплазме, и, следовательно, там же и происходит транскрипция их генома. Мы уже теперь хорошо знаем, что для транскрипции ДНК (иными словами, для синтеза иРНК на матрице ДНК) нужен фермент РНК- полимераза, зависимая от ДНК. Вирусы охотно пользуются для этих целей клеточными ферментами, подставляя свои матрицы под действие энзима клетки. Но для этого необходимо, чтобы там, где происходит репродукция данного вируса, этот фермент был. В цитоплазме такого клеточного фермента нет и быть не может, это чисто ядерный фермент.
Как же происходит тогда транскрипция генома этих вирусов, репродуцирующихся в цитоплазме? Одним единственно возможным способом, за счет активности фермента ДНК-зависимой РНК-полимеразы, содержащегося в структуре самого вириона (вирионной транскриптазы). Другие ДНК-содержащие вирусы (папова, адено, герпес, парво, гепадно), репродуцирующиеся в ядрах, транскрибируют свой геном за счет ядерных транскриптаз (ДНК- зависимых РНК-полимераз) клетки.
Так по-разному своими собственными, подчас неповторимыми путями вирусы транскрибируют свои геномы, и в клетках появляются вирусспецифические иРНК, способные транслировать в клеточных рибосомах вирусные белки.
Мы помним, что трансляцией называют процесс перевода генетической информации, содержащейся в иРНК, в специфическую последовательность аминокислот. Сама по себе трансляция - сложный, но достаточно хорошо изученный процесс. Известно, что происходит она в рибосомах (и только в них!) и проходит в три фазы: инициации, элонгации и терминации.
Не будем их описывать, это сделано в учебниках по молекулярной биологии. Нам важно другое: выяснить, как реализуется стратегия вирусного генома при трансляции в клеточных рибосомах вирусных белков.
У вируса должна быть своя стратегия, причем весьма "хитроумная", так как количество вирусных иРНК всегда значительно уступает количеству клеточных, и, следовательно, недостаточно просто наготовить свои молекулы, надо еще избирательно подавить трансляцию клеточных иРНК. Чтобы это сделать, надо хорошо "знать", как вообще начинается (инициируется) трансляция в рибосомах, и затем использовать это "знание" для своих целей. Стратегия вирусного генома как раз все это и предусматривает.
Инициация вообще является наиболее ответственным этапом трансляции. Именно в этот период рибосома должна "узнать" иРНК и связаться с ее особым участком. Трансляция начинается, когда к комплексу иРНК с малой рибосомальной субъединицей присоединяются так называемые инициаторные белки. Их по крайней мере три - в прокариотических (не обладающих типичным клеточным ядром и хромосомным аппаратом) клетках и более девяти в эукариотических (у которых тело клетки дифференцировано на цитоплазму и ядро). Таким образом, в каждой клетке имеется как бы пульт управления процессами трансляции: все иРНК, как правило, находятся в избытке, а инициаторные белки регулируют, какой вид трансляции и когда должен протекать.
Вирус вводит свои вирусспецифические инициирующие факторы, которые проводят "дискриминационную политику", создавая условия для избирательной трансляции вирусных иРНК. Трансляция клеточных иРНК полностью или частично прекращается, и тогда клетка более или менее быстро может погибнуть, либо совсем не подавляется, и тогда клетка продолжает жить и функционировать, несмотря на вирусную инфекцию. Но в любом варианте трансляция вирусных белков имеет режим наибольшего благоприятствования, и биосинтез структур чуждого для клетки организма, "тела" ее врага, клеточные рибосомы исправно осуществляют. Вот какова стратегия вирусного паразитизма!
Мы не будем говорить о деталях процесса трансляции вирусных белков. В одних случаях синтезируется один гигантский белок - предшественник, который потом нарезается на отдельные белки зрелого вириона (ведь у любых вирусов индивидуальных белков бывает много: от трех до нескольких десятков); в других - изначально раздельно синтезируются индивидуальные белковые молекулы. Почти во всех случаях белки подвергаются так называемой посттрансляционной модификации: изменяются за счет включения в их состав углеводов, сульфатных групп, молекул жирных кислот и т. д. Это тоже элементы стратегии вирусного генома, но лишь элементы. Главная задача вируса состоит в том, чтобы захватить клеточные рибосомы и заставить работать на себя. С этой задачей вирусный геном успешно справляется, сохраняя свой собственный вид и ставя под угрозу или попросту уничтожая клетки хозяина.
Итак, в итоге транскрипции и трансляции созданы вирусные белки. Теперь остается рассмотреть, как происходит синтез молекул нуклеиновой кислоты, гомологичных геному, чтобы в общих чертах понять финал драмы вирус - клетка. Синтез таких молекул носит название репликации.
Строго говоря, этот термин не является синонимом слов размножение и репродукция. Хотя нередко можно прочитать или услышать: "вирусы реплицируются" (при этом имеется в виду, что они размножаются). Но точнее, по существу, репликацией следует называть именно синтез новых, гомологичных исходной молекул нуклеиновой кислоты.
Из описания жизни клетки-государства мы должны помнить, что в клетке происходит репликация ДНК, в результате чего образуются дочерние двунитевые ДНК - знаменитые двойные спирали Уотсона и Крика. Как происходит этот процесс в клетке - известно достаточно хорошо, но так ли реплицируется вирусный геном?
Если у вируса двуцепочечная ДНК, а таких большинство (это и папова-, и адено-, и герпесвирусы и семейство вирусов оспы), то механизм репликации просто идентичен такому в клетке. С той лишь, однако, существенной оговоркой, что реплицируют ДНК вируса клеточные ферменты. А в остальном все точно так же: полинуклеотидные цепи двойной спирали ДНК расплетаются, и образуются две новые двойные спирали, каждая из них состоит из одной родительской и одной вновь синтезированной цепи.
Ученым понадобились многие десятилетия, чтобы раскрыть эту тайну жизни, тайну размножения, тайну передачи наследственных свойств. Через наивные, а подчас и мистические представления о природе похожести детей на родителей, через эмпирические наблюдения и строгие, но частные эксперименты, человеческий ум совсем недавно постиг тайну тайн живой природы, секрет двойной спирали ДНК и ее репликации, лежащей в основе всех видов размножения.
Вирус превзошел эту премудрость многие миллионы лет назад, научившись жить в живых клетках. Мастерски подставляя свой ДНК-овый геном под действие комплекса клеточных ДНК-синтезирующих ферментов, он теми же способами, теми же механизмами достигает собственных целей, реплицирует свой геном, продолжает жизнь своего биологического вида. Обманутая им клетка, может, мы уже упоминали об этом, и сразу погибнуть, и долго болеть, погибая постепенно, и вовсе не погибать, "делая вид", что ее не касается то, что в ней самой происходит. Именно так возникают латентные и хронические инфекции, в которых вирус или вирусный геном персистируют. Это тоже, конечно, репродукция с репликацией генома, но качественно иная, как бы растянутая во времени, как при замедленной киносъемке...
Впрочем, все это относится ко всем вирусам вообще, а мы говорим пока о репликации генома ДНК-содержащих вирусов. Их геномы реплицируются так, как ДНК обычных клеток. Есть, конечно, особенности, но они носят частный характер. Так, у некоторых ДНК-содержащих вирусов геном представляет собой кольцевую двунитевую структуру (например, у вирусов группы папова). В этом случае ферменты разрезают одну из нитей ДНК, это приводит к раскручиванию молекулы, снятию так называемых супервитков, и возникает обычная (точнее - типичная) линейная структура ДНК, а далее - все как у Уотсона и Крика, так называемым полуконсервативным способом, суть которого описана выше.
Есть вирусы, правда, их меньшинство, с однонитевым ДНК-овым геном (семейство парвовирусов). Но и эти, казалось бы, "инвалиды", поступают типично, хотя и с ухищрениями. До репликации на матрице единственной цепи ДНК генома этих вирусов с помощью опять-таки клеточных (!) ферментов строится комплементарная ей нить ДНК, и возникает типичная, хотя и временная, двойная спираль. Это сооружение недолговечно, оно служит только для репликации, из него выходят вирусы, содержащие только одну нить ДНК (как и их "родители"). Структуры эти носят название репликативных форм. Весьма примечательно, что хоть на время, но возникают стандартные двуспиральные комплексы, в которых идет репликация. Природа во всем своем разнообразии опирается на оптимальные решения и конструкции. Двойная спираль является одним из таких оптимумов.
Все, что говорилось до сих пор, касалось ДНК-содержащих вирусов. Ну а как реплицируются геномы их РНК-содержащих собратьев? В этом случае - все так и все иначе. В клетке нет и не может быть ферментов, способных реплицировать РНК. Поэтому все ферменты, участвующие в этом процессе, - вирусспецифические. В основном это те же транскриптазы, только модифицированные...
Мы уже не раз говорили, как разнообразно подцарство рибовирусов: сколь различен геном у разных их представителей, как по-разному идет у них транскрипция и трансляция. Нет идентичности, конечно же, и в процессе умножения их генетического материала, то есть репликации. Но мы надеемся, что теперь-то читатель нам поверит: при всем разнообразии деталей - итог один - масса геномной РНК в инфицированной клетке увеличивается, число копий геномной РНК резко возрастает, а сама репликация проходит все в тех же излюбленных природой формах, в репликативных комплексах (только на этот раз РНК→РНК).
Итак, мы почти дошли до финала. В клетке, пораженной вирусами, наработаны порознь (в разных структурах клетки) вирусные нуклеиновые кислоты и вирусные белки. Вирусные нуклеиновые кислоты синтезируются в ассоциации с клеточными структурами, которые расположены вдали от рибосом, где происходит наработка вирусных белков. Им предстоит еще найти друг друга, узнать и объединиться, чтобы стать вирусом.
Ничего подобного природа не знает! Наивные представления средневековых схоластов о гомункулюсе, маленьком человечке, все внутриутробное развитие которого сводится к тому, что он растет и становится все больше и больше, можно если не понять, то по крайней мере оправдать: никто ведь не видит своими глазами эмбрион как таковой! Но кому может прийти в голову, что при размножении голова образуется где-то в одном месте, ноги в другом, а туловище - в третьем, а потом это все собирается в одно тело?
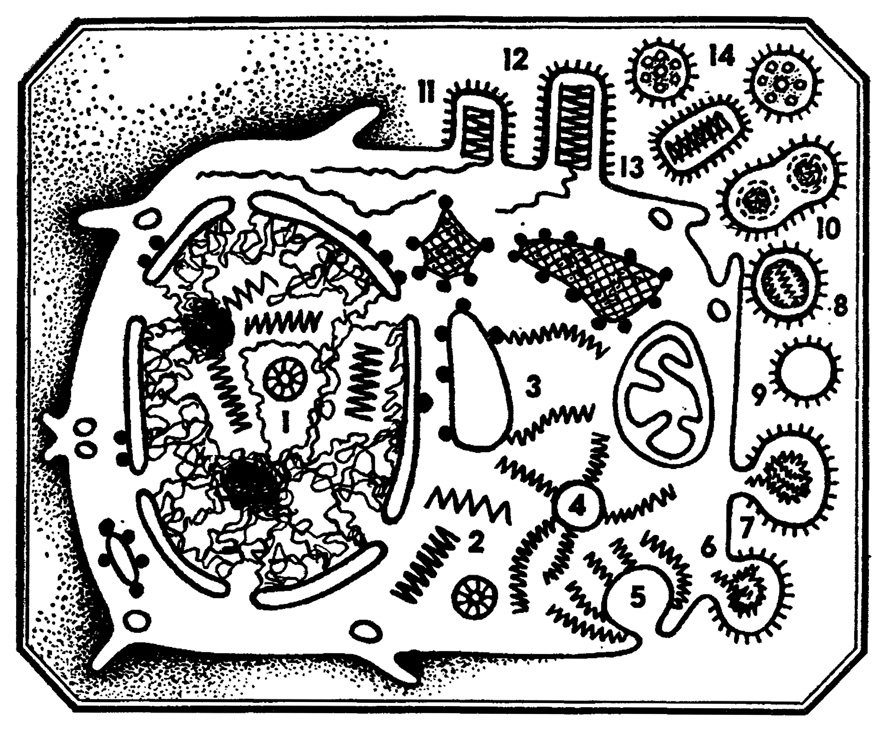
Рис. 6. Морфогенез вируса гриппа. 1 - скопление нуклеокапсидов вируса в нуклеоплазме; 2 - скопление нуклеокапсидов вируса в гиалоплазме; 3, 4 - этапы формирования вирусного рибонуклеопротеида; 5 - 7 - предполагаемые этапы формирования овальных вирионов на поверхности клетки; 8 - внеклеточный вирион; 9 - внеклеточный неполный вирус; 10 - внеклеточный нуклеокапсидный вирион; 11, 12 - этапы формирования цилиндрических вирионов на поверхности клетки; 13, 14 - внеклеточные цилиндрические вирионы в различной ориентации
Но вирусная самосборка происходит именно так: вирусные нуклеиновые кислоты и белки при достаточной концентрации обладают способностью узнавать друг друга в многообразии клеточных белков и нуклеиновых кислот и соединяться друг с другом. Это и есть самосборка! Конечно, бывают и сбои: где-то осталась голая нуклеиновая кислота, где-то белок не нафаршировался нуклеиновой начинкой, но это все брак, его, как правило, немного. Основная масса компонентов собирается правильно, и полчища зрелых вирионов покидают клетку либо путем взрыва, либо путем почкования.
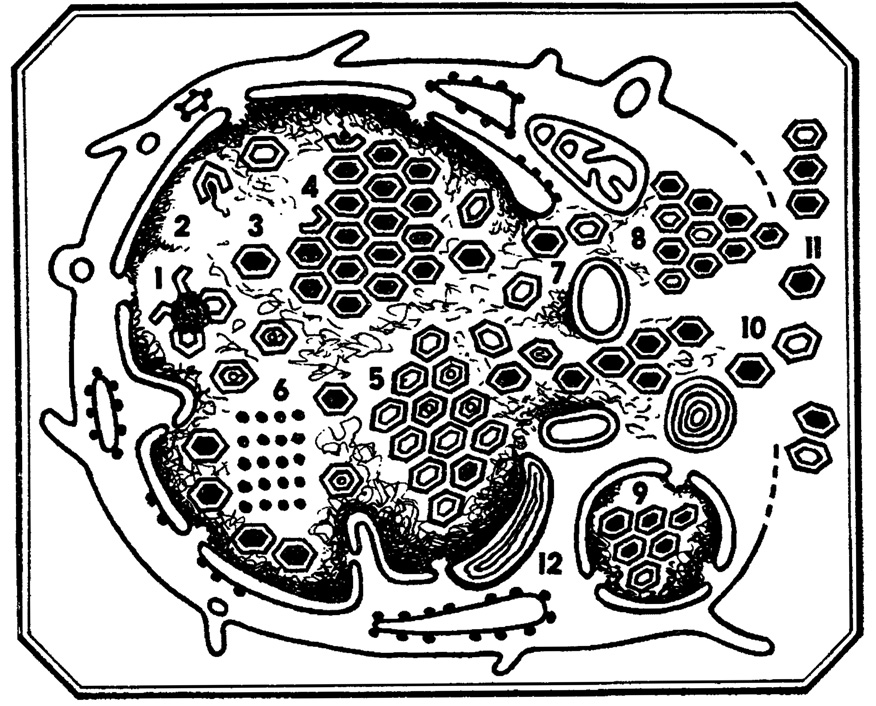
Рис. 7. Внутриклеточный морфогенез аденовирусов. 1, 2 - формирование нуклеокапсидов; 3 - вирион; 4 - кристаллоподобное внутриядерное скопление полных вирионов; 5 - кристаллоподобное цитоплазматическое скопление вирионов; вирионов; 6 - пара-кристаллическое белковое скопление; 7 - выделение вирионов из ядра в протоплазму через расширенные поры ядерной оболочки; 8 - кристаллоподобное цитоплазматическое скопление вирионов; 9 - скопление вирионов в участке ядра, отпочковавшегося в результате фрагментации ядра клетки; 10 - выделение вирионов из клетки; 11 - внеклеточные вирионы; 12 - участок редуплицирующейся ядерной оболочки
Взрыв равносилен полной цитодеструкции, разрушению клетки, в результате чего вирусные частицы оказываются в окружающей среде. Так расправляются со своей alma mater (от латинского мать вскормившая, образно: родной дом) пикорнавирусы, аденовирусы и другие агенты, не имеющие липопротеидной оболочки поверхностного белкового капсида. Почкование характеризует выход из клеток вирусов, содержащих липопротеидную мембрану (например, орто- и парамиксовирусы), которая является дериватом клеточных мембран. Итак, обманув "правительство" клетки-государства, создав свое потомство с помощью ее механизмов, эти вирусы напоследок воруют куски клеточных оболочек. Несмотря на такой разбой, клетки могут еще какое-то время продуцировать новые полчища своих губителей, но в конце концов все-таки разрушаются...
Драма окончена: вирус репродуцировался, клетка погибла! Занавес!
Пока ученые не разобрались в механизме процесса репродукции, им мнилось всякое, в том числе сверхъестественное. Судите сами. Сквозь оболочку клетки проникает вирион: вот он, виден в микроскоп, определяется другими методами. Сомнений в его присутствии нет. Но через короткий промежуток времени никаких доказательств его наличия в природе вообще нет, самые точные и совершенные методы обнаружить его в клетке не могут. Исчез. Растворился. Клетка его переварила... И вдруг описанная выше драма. Оказывается, погиб не вирус, а сама клетка. Вирион же, как сказочная птица феникс, возродился из пепла, а точнее из ничего. Да не один, а "со товарищи".
Долго не могли понять и другое, почему всегда все вирионы одного возраста. Почему среди вирусов (или у вирусов, если все же считать их существами) нет юношей, нет стариков, все одинаково зрелые. Лишь в последние десятилетия, изучив механизм репродукции, ученые нашли вполне материалистические ответы на "мистические" вопросы.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://nplit.ru/ 'Библиотека юного исследователя'