ЧАСТЬ III: СПЕЦИАЛЬНЫЕ ГЛАВЫ
ГЛАВА 8: ИСТОРИЯ ЖИЗНИ НА ЗЕМЛЕ
Происхождение жизни, первичные микроорганизмы. Древнейшие следы жизни. Эволюция растений. Происхождение типов животных. История членистоногих. История позвоночных животных - рыб, земноводных, пресмыкающихся, птиц, млекопитающих. Эволюция биомассы. Эволюция форм организмов, мутации и адаптация. Живые ископаемые. Гигантизм. Вымирание. Миграции. Происхождение человека
Если бы стало известно, что на какой-то планете есть жизнь, то первым вопросом о ней был бы вопрос: «Какая жизнь?». Ответ для Земли: белков о-нуклеинокислотная. Второй вопрос - «Сколько ее?» (т. е. какова суммарная биомасса). Ответ для Земли: сейчас 2-3 триллиона т. Третий и четвертый вопросы - «Каковы формы живых организмов? Какие из них преобладают?». Ответы для Земли: 35 типов наземных и водных растений и животных; из суммарной биомассы сейчас 98-99% находится на суше, в основном это растительность лесов, и 34 млрд. т - в океане (из них две трети - зоопланктон, около трети - донные животные, 5% - водоросли и 3%, или 1 млрд. т, - рыбы и кальмары). Можно было бы задавать еще много разных вопросов, но в этой книге хочется отдать первое место следующему: как менялись в течение истории Земли формы и количества живых организмов? Ответу на этот вопрос мы и посвящаем настоящую главу.
При решении вопроса о происхождении жизни на Земле прибегать к гипотезе о панспермии, т. е. о переносе «семян жизни» с одних планет на другие и даже из одних планетных систем в другие, нам казалось бы попыткой уйти от поисков естественных процессов развития высокоорганизованной материи. Изложение возможных процессов образования живого из неживого потребовало бы специальной книги. Мы имеем возможность отослать читателя к книгам А. И. Опарина [45] и А. И. Опарина и В. Г. Фесенкова [46], а также к обсуждению вопроса о сущности жизни в книгах Э. С. Бауэра [47], Э. Шредингера [48] и И. С. Шкловского [49], ограничжваявь дало лишь несколькими короткими формулировками.
Жизнь на Земле зародилась, когда на дне мелких теплых морей катархея, богатых сложными органическими веществами до аминокислот включительно, начали образовываться двойные сахаро-фосфатные спиральные нити высокополимерных нуклеиновых кислот с закрепленными на них последовательностями оснований (служащими «кодами» для синтеза белков), способные при некоторых условиях разворачиваться в одинарные спирали и синтезировать на каждой из них недостающую вторую спираль, т. е. порождать пару себе подобных (и передавать им информацию о процедурах синтеза белков, закодированных последовательностями оснований). Такие полимеры, которые синтезировали белки, обеспечивающие им достаточно длительное самосохранение, по-видимому, уже можно считать первичными организмами.
Эти первичные микроорганизмы, возможно, питались имевшейся органикой небиологического происхождения, осуществляя, например, бескислородное разложение белков и аминокислот - гниение или углеродов - брожение (анологично питаются современные сапрофиты, поглощая через свои стенки клеток органику биологического происхождения: например, дрожжи при отсутствии кислорода сбраживают глюкозу до спирта и углекислоты). Развитие этих организмов, вероятно, исключило условия для дальнейшего самозарождения жизни, и с тех пор все живое появляется уже только от живого. В результате эволюции микроорганизмов (вынуждаемой нехваткой подходившей для пищи органики) у них появилась способность синтезировать необходимые для самосохранения органические молекулы из неорганических. Наиболее эффективным способом оказался фотосинтез - продуцирование органического вещества из углекислоты и воды под действием солнечного света (энергия света, поглощаемого пигментами, прежде всего зеленым хлорофиллом, расходуется на расщепление молекул воды, кислород выделяется в атмосферу, а водород вместе с углекислотой идет на образование первичного органического продукта - фосфоглицериновой кислоты).
Первыми фотосинтезирующими растениями были, по-видимому, микроскопические синезеленые водоросли цианофиты, у которых хлорофилл рассеян в виде мелких зерен по плазме клеток (у более развитых растений он сосредоточен в специальных тельцах - хлоропластах), и имеется еще синий пигмент фикоцианин. Эти водоросли похожи на бактерий тем, что в их клетках трудно различить ядра, и размножаются они только делением.
Упоминавшиеся в главе 5 наиболее древние остатки жизнедеятельности организмов, найденные в Трансваале в породах серии Свазиленд возрастом 3.1-3.4 млрд. лет, были тщательно изучены Э. Баргхорном и Дж. Шопфом. Они представляют собой микроскопические изолированные палочки длиной 0.45-0.7 мк и диаметром 0.18-0.32 мк, имеющие двухслойные оболочки толщиной 0.045 мк; там же обнаружены нитеподоб-ные образования, а также микроскопические шаровидные, дисковидные и многоугольные оболочки одноклеточных водорослей акритархи. Почти столь же древними (более 2.9 млрд лет) являются обызвествленные продукты жизнедеятельности цианофитов и бактерий - прикрепленные ко дну столбчатые строматолиты и неприкрепленные округлые онколиты, найденные в известковых прослоях зеленокаменных пород системы Булавайо в Южной Родезии и описанные А. Мак-Грегором еще в 1940 г. (назовем также узорчатые карбонатные микростяжения - катаграфии).
Упомянем еще строматолиты возрастом 1.9 ± 0.2 млрд. лет в кремнистых сланцах формации Ганфлинт канадской провинции Онтарио, где найдены и обстоятельно изучены Э. Баргхорном и С. Тайлером (1965 г.) многочисленные нитеподобные, звездчатые, округлые и более сложные «микроорганизмы», содержащие органическое вещество. В более молодых осадочных породах строматолиты, онколиты и катаграфии находятся во все возрастающих количествах. В рифее этих форм уже так много, что по их комплексам можно различать породы нижнего, среднего и верхнего рифея и венда (М. А. Семихатов [17]).
В главе 5 уже упоминалось об одном из наиболее древних следов животных, найденном А. М. Лейтесом [50] в среднепротерозойских слоях Удоканской серии Забайкалья. Он представляет собой приуроченные к строматолитовым породам тонкостенные трубочки длиной до 2.5 см, диаметром до 4 мм, имеющие четырехгранное сечение с округленными углами и оставшиеся, по-видимому, от червей (если это так, то кишечнополостные животные должны были существовать и еще раньше). По-видимому, к началу рифея кислорода было уже достаточно для развития примитивных животных. В породах среднего и верхнего рифея найдены следы роющих животных и трубочки червей. В породах венда обнаружены уже многочисленные отпечатки разнообразных бесскелетных животных, а начало кембрия маркируется массовым появлением скелетообразующих организмов.
Палеонтологические данные позволили воссоздать грандиозную и стройную картину эволюции форм жизни в фанерозое (рис. 37), не оставляющую никаких сомнений в естественности этого процесса. В предыдущей главе мы «вырезали» из этой картины поочередно каждый геологический период и давали очень краткую характеристику животного и растительного мира Земли в соответствующий промежуток времени. Теперь же рассмотрим рис. 37 не по строчкам, а по столбцам, т. е. проследим фанерозойскую эволюцию ряда важнейших типов организмов.

Рис. 37. Распространение типов организмов в различные периоды фанерозоя.
Начнем с растений, которые осуществляют фотосинтез первичного органического вещества и потому являются первоосновой всей жизни. Водоросли появились еще в начале архея (или, может быть, даже в катархее) и с тех пор благополучно существуют до настоящего времени преимущественно в виде одноклеточных форм. Доля водорослей в общей массе живых организмов на Земле в наше время составляет меньше одной тысячной (около 1.7 млрд. т из общей суммы в 2-3 триллиона т), но зато они чрезвычайно быстро размножаются - их клетки делятся каждые 1-2 суток, и в результате их продукция, около 550 млрд. т в год, составляет основную долю продукции биомассы на Земле в целом.
Первые наземные растения появились, вероятно, в ордовике или даже в кембрии, но достоверные их следы найдены лишь в силуре. Это были самые примитивные из высших расте; ний типа сосудистых - псилофиты, мелкие растения без корней и листьев, а также первые плауны. В девоне появились представители двух других подтипов сосудистых растении - клинолистных (хвощей) и папоротникообразных (из последних - пока лишь наиболее примитивные растения класса папоротников а также отряда семенных папоротников птеридосперм из класса голосемянная), причем крупные древовидные формы плаунов хвощей и папоротников образовали первые леса нашей планеты. В карбоне обширные участки суши покрыли огромные заболоченные леса из гигантских плаунов лепидодендронов и сигиллярий, хвощей каламитов, древовидных папоротников и птеридосперм, а также новых отрядов вечнозеленых голосемянных кордаитов, а к концу периода - первых хвойных растении и гинкго. В перми появились бенеттиты, плауны пришли в упадок; на территориях Гондваны господствовала специфическая растительность во главе с голосемянными глоссоптерисами.
Триас и юра - это время расцвета голосемянных растений отрядов бенеттитов, хвойных, гинкго и цикадовых (саговников); птеридоспермы и кордаиты вымерли. В нижней половине мела, а возможно даже в юре, появились первые покрытосемянные (цветковые) растения, сначала двудольные. Во второй половине мела появились также однодольные цветковые растения, а двудольные образовали уже леса (кленовые и дубовые). Они вытеснили ряд отрядов голосемянных - бенеттиты вымерли, цикадовые и гинкго пришли в упадок. Наступивший затем кайнозой - эра расцвета цветковых растений.
Рассмотренная эволюция форм растений от водорослей до цветковых ттооисходила по линии уменьшения роли двуполых гаплоидных поколении, в телах которых (гаметофитах) клетки содержат одинарные наборы хромосом, и увеличений роли бесполых диплоидных поколений, в телах которых клетки содержат двойные наборы хромосом. Так, например, у многоклеточной водоросли улотрикс имеется только одна диплоидная клетка, а все остальные гаплоидны. У покрытосемянных, наоборот, все растение гаплоидно а гаметофит состоит лишь из нескольких клеток в тканях цветка; эволюционная выгода этого заключается как в том, что для размножения цветковых не требуется внешней влаги, так и в меньшей чувствительности диплоидных организмов к влиянию рецессивных генов (таких, которые действуют лишь при наличии их в обоих наборах хромосом в диплоидной клетке).
В наше время наземные растения, преимущественно цветковые и хвойные, составляют 98-99% от 2-3 триллионов т биомассы на Земле, но растут они медленно - их годичная продукция составляет всего около 50 млрд. т (вдесятеро меньше продукции водорослей в океане), а средний возраст (очевидно, возраст деревьев в лесах) - 40-60 лет.
Переходя к эволюции животных, отметим, что сведения палеонтологии о времени появления, расцвета и вымирания тех или иных групп животных вместе с данными о степени сложности их устройства, способах размножения и особенностях развития их зародышей позволяют судить о происхождении этих групп. Современные представления о происхождении основных типов животных (базирующиеся на принципе монофилии, т. е. на предположении о наличии единого корня происхождения каждой группы организмов) показаны на рис. 38. Согласно этим представлениям, одноклеточные простейшие животные породили как тупиковую ветвь губок, так и прогрессивную ветвь кишечнополостных, от которой пошли два главных ствола - первичноротых и вторичноротых. На первом из этих стволов промежуточным этапом были низшие черви, от которых пошли ветви немертин, плеченогих, мшанок, форонид, моллюсков и через кольчатых червей, наконец, членистоногие. От другого ствола поочередно отделялись ветви щетинкочелюстных червей, иглокожих, погонофор, кишечнодышащих, оболочников, головохордовых и, наконец, позвоночных.
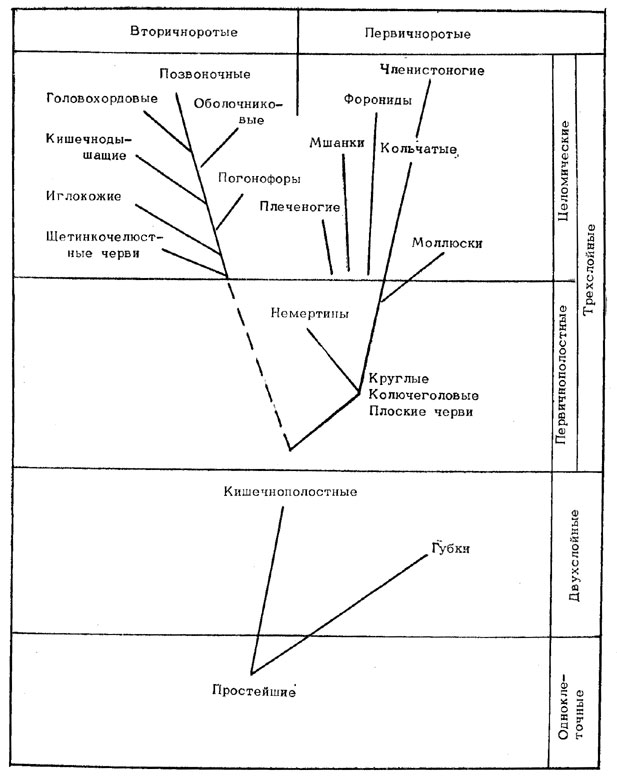
Рис. 38. Происхождение основных типов животных.
Ряд типов животных - по крайней мере простейшие, кишечнополостные и некоторые низшие черви - появился, хотя бы в небольших популяциях, еще в среднем протерозое. В качестве примера фанерозойской эволюции простейших на рис. 39 приведен график числа семейств фораминифер (появившихся в самом начале кембрия). Число семейств - одна из возможных количественных характеристик разнообразия форм данного типа организмов, свидетельствующая о темпах его эволюции: в периоды быстрой эволюции разнообразие форм должно возрастать. На графике видно, что для фораминифер в палеозое наиболее динамичными были периоды расцвета фузулинид - карбон и особенно пермь, триас оказался периодом застоя и спада, в юре разнообразие вновь достигло пермо-карбонового уровня, в течение мела оно резко возрастало и достигло нового максимума в эпоху нуммулитов - в палеогене.
![Рис. 39. Изменение числа семейств фораминифер в фанерозое по Г. П. Леонову [41].](pic/000049.jpg)
Рис. 39. Изменение числа семейств фораминифер в фанерозое по Г. П. Леонову [41].
К сожалению, мы не имеем возможности остановиться на истории развития каждого из типов животных - напомним, что всего имеется 23 типа! Поэтому о первичноротых мы скажем лишь то, что все их типы появились, по-видимому, еще в докембрии. Нам придется оставить в стороне такие интересные и важные вопросы, как, например, связанная с эволюцией кишечно-полостных история коралловых рифов или же охватывающая весь фанерозой история «приматов моря» - головоногих моллюсков (не раз порождавших и массовые формы, и таких гигантов, как наутилусы ордовика с 6-метровыми прямыми раковинами, меловые аммониты со спиральными раковинами 3-метрового диаметра и современные гигантские «кракены» - глубоководные кальмары архитевтисы длиной до 15-20 м, а может быть, и еще больше). Остановимся лишь на членистоногих, ставших вершиной эволюции первичноротых животных.
Уже в кембрии сформировался подтип наиболее ранних и примитивных членистоногих - трилобиты (рис. 31). В кембрии и ордовике они буквально кишели на дне прибрежной зоны морей и океанов; в силуре их количество начало убывать, и к концу палеозоя они вымерли. В кембрии уже существовали и представители подтипа жабродышащих (класса ракообразных).
Так же рано появились первые членистоногие подтипа хелицеровых (т. е. клешнеусых) - огромные хищные ракоскорпионы эвриптериды, или гигантостраки, достигавшие в длину иногда 2 м, - самые крупные из всех членистоногих, когда-либо живших на нашей планете. Они просуществовали почти весь палеозой. Еще в силуре от них произошли первые сухопутные (дышащие воздухом) животные, похожие на современных скорпионов, у которых брюшные жаберные ножки эвриптерид превратились в легкие. Но такой орган воздушного дыхания оказался для членистоногих не оптимальным; в девоне от кольчатых червей произошли сухопутные членистоногие подтипа трахейнодышащих, в том числе класса насекомых, которые быстро завоевали сушу и настолько преуспели, что уже в карбоне появились крылатые насекомые, включая гигантских стрекоз палеодиктиоптер с размахом крыльев до 90 см. Членистоногие в наши дни представляют собой самый динамичный, т. е. богатый формами, тип животных: из описанных учеными 1.5 млн. видов животных более 1 млн. приходится на членистоногих (и из них более 90 % - на насекомых). Они составляют немалую долю и от массы животных на Земле: так, например, суммарная масса океанического зоопланктона, состоящая преимущественно из ракообразных, равняется 21.5 млрд. т - около двух третей всей биомассы Мирового океана.
Из вторичноротых рассмотрим лишь историю позвоночных. Первые их представители появились в ордовике (а может быть, даже еще в кембрии), это так называемые панцирные рыбы - небольшие рыбообразные донные животные без челюстей и плавников, покрытые панцирем из костных пластинок на голове и толстых чешуи на туловище; они еще не были настоящими рыбами - из современных позвоночных животных к ним ближе всего бесчерепные (ланцетники) и бесчелюстные - круглоротые (миноги и миксины). В силуре появились, а в девоне получили широкое разбитие уже и настоящие рыбы, как хрящевые (сначала панцирные, а затем и беспанцирные акулы, благоденствующие с тех пор по настоящее время), так и костные, уже в середине девона поделившиеся на лопастеперых (двоякодышащих и кистеперых) и лучеперых, сначала ганоидных, особенно процветавших в перми и триасе, а с юры и костистых, имеющих абсолютное господство среди рыб в настоящее время.
Девон был временем первых попыток рыб выбраться на сушу; двоякодышащие рыбы в этом не преуспели, а от кистеперых пошли земноводные животные (амфибии), которые осваивали сушу одновременно с насекомыми. Этих древних амфибий называют стегоцефалами, т. е. панцирноголовыми, так как их голова (а у некоторых и брюшная сторона) была покрыта защитными костными пластинками. Крупных стегоцефалов карбона и перми называют лабиринтодонтами по петлеобразному эмалевому рисунку на поперечном разрезе их зубов (который был свойственным еще и кистеперым рыбам). Большинство их вымерло в перми, оставшиеся - в триасе; в юре сформировались современные бесхвостые амфибии (лягушки и т. п.), в мелу - хвостатые (тритоны и саламандры) и уже в кайнозое - безногие (червяги).
Первые настоящие наземные позвоночные - это пресмыкающиеся (рептилии), у которых размножение и развитие происходят на суше. История рептилий перми и мезозоя, этих двухсот миллионов лет господства чудовищ на нашей планете, является, вероятно, самой драматической главой истории жизни на Земле до появления человека. Первые рептилии появились в верхнем карбоне; это были представители подкласса анапсид, имеющих цельный черепной панцирь, - небольшие (до полуметра длиной), очень похожие на стегоцефалов и просуществовавшие только в течение перми сеймурии и более крупные котилозавры, существовавшие до конца триаса (еще в перми от них произошли черепахи). В конце карбона от котилозавров пошли рептилии подкласса синапсид с черепным панцирем, сократившимся путем образования одной височной ямы, ограниченной снизу скуловой дугой, что усилило челюсти. В конце карбона и ранней перми это были стройные звероподобные хищники пеликозавры, со средней перми - зверообразные тероморфы, вымершие в начале мезозоя (в их числе крупнейший верхнепермский хищник - 3-метровая саблезубая иностранцевия). Также, по-видимому, от котилозавров пошли водные рептилии - во-первых, подкласса синаптозавров, еще сохранивших в какой-то мере облик наземных животных (в том числе длинношеие ластоногие плезиозавры, длиной от полуметра до 15 м, существовавшие с позднего триаса до конца мела), и, во-вторых, подкласса ихтиоптеригий (сначала небольшие мезозавры, а со среднего триаса - огромные рыбообразные ихтиозавры, достигавшие в длину до 18 м, которые вымерли в конце мела).
Основными же мезозойскими рептилиями были диапсиды, у которых черенной нанцнрь стегоцефалов сократился нутем образования двух височных ям, ограниченных снизу скуловыми дугами. Из диапсид подкласса лепидозавров назовем, во-первых, известных с раннего триаса клювоголовых (от которых ныне осталась только гаттерия островов района Новой Зеландии); во-вторых, существовавших в верхней перми ящерицеобразных эозухий; в-третьих, существующих с конца юры и до сегодняшнего времени настоящих ящериц - чешуйчатых (от которых в начале мела пошла ветвь морских ящеров мозазавров длиной до 12 м, просуществовавших до конца мела, а в конце мела произошли змеи). Наиболее широкое развитие получили диапсиды подкласса архозавров. Первыми из них были происшедшие от эозухий и появившиеся в начале триаса небольшие животные псевдозухии с относительно длинными задними ногами. От них пошли, во-первых, крокодилообразные водные фитозавры триаса, породившие в юре настоящих крокодилов, которые в течение мела уже приняли современный облик; во-вторых, летающие ящеры юры и мела птерозавры (от совсем маленьких - с воробья - до мелового птеранодона с размахом крыльев, достигавшим 8 м) и, в-третьих, птицетазовые и ящеротазовые динозавры.
Особенно многообразными были растительноядные птицетазовые динозавры, в том числе, например, передвигавшиеся на трехпалых задних лапах игуанодоны высотой до 5-9 м, четвероногие стегозавры 10-метровой длины с двойным гребнем метровых треугольных костных пластин на спине, трехрогий меловой трицератопс с широким костяным «воротником». В числе же ящеротазовых были как растительноядные четвероногие, достигавшие колоссальных размеров (бронтозавры длиной до 20 м и весом, вероятно, до 30 т, диплодоки длиной свыше 25 м и самые огромные из всех сухопутных животных, когда-либо проживавших на Земле, - брахиозавры длиной около 24 м и весом, должно быть, порядка 50 т), так и хищники, которые передвигались на задних лапах, опираясь на хвост (как ныне кенгуру), причем некоторые из них достигали огромных размеров (например, 15-метровый меловой тираннозавр реке, крупнейший из наземных хищников за всю историю жизни на Земле).
Современное представление о пресмыкающихся,- как о холоднокровных животных с чешуйчатой кожей, плохо приспособленных к среде с низкой или резко колеблющейся температурой, возможно, не следует распространять на динозавров. Для выяснения этого вопроса Роберт Беккер (1975 г.) использовал три признака, отличающих теплокровных животных от холоднокровных:
1) наличие в костях большого количества кровеносных сосудов, а также каналов, в которых происходит быстрый обмен фосфатом кальция, необходимый для нормального функционирования мышц и нервов;
2) малое отношение биомасс хищников и их жертв (порядка 1-5% против десятков процентов у холоднокровных животных);
3) способность существовать в условиях холодного климата.
По всем трем признакам установлено, что если раннепермские хищные пеликозавры (например, диметродоны) еще были холоднокровными, то уже позднепермские терапсиды и позднетриасовые текодонты были теплокровными (отношения хищник - жертва порядка 10-15%); у динозавров признаки теплокровности выражены даже ярче, чем у современных млекопитающих (отношение хищник-жертва всего 1-3%, так что находки костей хищных динозавров чрезвычайно редки). Динозавры, особенно жившие в умеренных и высоких широтах, вероятно, имели и волосяные покровы.
И все это царство чудовищ полностью вымерло в конце мелового периода! Правда, среди современных рептилий тоже имеются опасные хищники, такие, как 6-метровые крокодилы, огромные ядовитые змеи - королевские кобры Юго-Восточной Азии (длиной до 5.5 м), африканские черные мамбы (более 4 м), южноамериканские бушмейстеры (более 3.5 м), австралийские тайпаны (до 3.5 м) и, наконец, «драконы острова Комодо», гигантские вараны длиной более 3 м, охотящиеся на оленей. Но все они не идут ни в какое сравнение с чудовищами, господствовавшими на Земле в мезозойскую эру.
От мелких динозавров, живших на деревьях, в конце юры произошли первоптицы - археоптериксы и археорнисы, которые были величиной с ворону, имели зубастые челюсти, слабые крылья с когтями на концах пальцев и длинные хвосты с многочисленными позвонками, покрытые перьями.
В породах мела найдены останки новых зубастых птиц - водяного гесперорниса и летающего ихтиорниса. В конце мела появились первые беззубые птицы. Современный облик они приобрели лишь в начале кайнозоя. Сейчас птицы играют очень большую роль в жизни нашей планеты - число особей этого класса оценивается в сотню миллиардов!
Млекопитающие имеют значительно более древнее происхождение, чем птицы: они возникли в триасе от палеозойских рептилий - каких-то мелких разновидностей тероморф отряда зверозубых. Такими первыми млекопитающими были просуществовавшие по палеоген многобугорчатые - растительноядные зверьки величиной с крысу, имевшие зубы с многочисленными бугорками и сильно развитые резцы, но не имевшие клыков. От их ранних форм, по-видимому, отошла ветвь однопроходных яйцекладущих млекопитающих (единственными сегодняшними представителями которых являются утконосы и ехидны Австралии, Тасмании и Новой Гвинеи), а в средней юре от них пошли многообразные трехбугорчатые (в частности, пантотерии) - мелкие зверьки, питавшиеся частью животной, а частью растительной пищей. От последних в мелу пошли и сумчатые, и плацентарные.
Первые сумчатые появились на материках северного полушария. Это были мелкие крысоподобные зверьки типа современных североамериканских опоссумов, имевшие бугорчаторежущие коренные зубы и крупные клыки. Затем они были вытеснены плацентарными в южное полушарие (в Северной Америке и на о. Сулавеси сохранилось лишь по одному виду) и дали толчок развитию многочисленных видов животных (так называемых сумчатых мышей, крыс, тушканчиков, кошек, медведей, волков, муравьедов, кротов, бандикутов, опоссумов, коал, вомбатов, валлаби и кенгуру) только в Австралии, куда из плацентарных проникли лишь летучие мыши и мелкие грызуны. В плейстоцене некоторые из кенгуру и вомбатов достигали размеров нескольких метров.
Первыми плацентарными были насекомоядные верхнего мела. От них произошли рукокрылые, древние хищники креодонты (в эоцене породившие зейглодонтов, от которых пошли китообразные, а на рубеже между эоценом и олигоценом разделившиеся на ластоногих и наземных хищников современных форм - куниц, вивер, собак и кошек), грызуны, неполнозубые, трубкозубые и приматы. Еще в палеоцене от креодонтов произошли первичные копытные - кондилятры, некрупные всеядные животные с бугорчатыми коренными зубами, умеренными клыками и пятипалыми конечностями с усиленным средним и ослабленными первым и пятым копытцами. От них произошли современные растительноядные парно- и непарнокопытные. В эоцене возникли и хоботные. Всего плацентарные млекопитающие образовали 31 отряд, из них 14 вымерли, а 17 существуют и в настоящее время.
История отряда приматов имеет для нас, конечно, особый интерес. Первые приматы - представители подотряда полуобезьян - произошли в палеоцене от древесных насекомоядных предков, внешне похожих на белок (такой облик сохранили самые примитивные из современных полуобезьян - тупайи). В эоцене от различных групп семейства наиболее развитых полуобезьян долгопятов произошли первые представители подотряда обезьян - широконосые обезьяны Центральной и Южной Америки, эволюционно оказавшиеся побочной ветвью, а через промежуточное звено парапитеков (остатки которых были найдены в нижнем олигоцене Египта) - узконосые обезьяны Старого Света, как низшие (собакоподобные), так и (через другие промежуточные звенья олигопитеков, проплиопитеков, эолопитеков и египтопитеков - нижний олигоцен северо-восточной Африки, так называемой Фаюмской депрессии) человекоподобные (гоминоиды). Последние сейчас делятся на три семейства - гиббоновых, с объемом мозга 100-150 см3, человекообразных обезьян, или понгид, с объемом мозга 350-600 см3 (орангутаны, шимпанзе и гориллы) и людей, или гоминид, с объемом мозга большим, чем у понгид (у современных людей - 1500 см3). Эти семейства произошли от различных миоценовых предков: гиббоны - от найденных в миоцене Африки лимнопишков, орангутаны - от индийских парасимий, а гориллы, шимпанзе и человек - от дриопитеков. К происхождению человека мы еще вернемся в конце этой главы.
Эволюция биомассы на Земле рисуется следующим образом. Разделим существующие организмы на группы:
1) водоросли,
2) морские животные,
3) наземные растения,
4) наземные животные. Водоросли появились в катархее, и в течение архея и протерозоя их биомасса нарастала до величины порядка сегодняшней, т. е. до 1 млрд. т; ее возможные колебания в фанерозое, вероятно, этого порядка величины не меняли. Морские животные появились в среднем протерозое, их биомасса очень медленно росла до венда, а в течение кембрия, вероятно, быстро выросла до величины порядка сегодняшней, т. е. до 20-30 млрд. т. Наземные растения появились, возможно, еще в ордовике или даже в кембрии, однако их биомасса стала ощутимой лишь в силуре, а за девон и карбон быстро выросла до колоссальных размеров, порядка нескольких триллионов тонн; в перми она, возможно, даже несколько уменьшилась и приблизилась к современному уровню. Наконец, наземные животные появились только в девоне, их биомасса достигла современной величины, вероятно, еще в палеозое.
Эти оценки изменений биомассы в течение истории Земли, конечно, чисто умозрительны (однако их дальнейшая детализация представляется не безнадежной). Что же касается изменений со временем разнообразия форм организмов, то здесь, наоборот, имеются довольно надежные фактические данные. Они показывают, что в разнообразии форм монотонного нарастания со временем не было, так как при появлении новых форм некоторые из старых отмирали (рис. 40, а). Так, в кембрии появилось 28 классов животных, в ордовике еще 14 (но часть классов уже отмерла), в силуре еще 3 (но 2 отмерли). Начало верхнего палеозоя ознаменовалось новой вспышкой формообразования - в девоне появилось 9 новых классов (и их число достигло фанерозойского максимума), а карбоне еще 5 (но 8 отмерло), в перми новых классов не появилось (но 4 отмерли). К триасу 3 класса отмерли, но 1 появился (и число классов достигло фанерозойского минимума, если не считать кембрия), в юре и мелу был небольшой прирост. В кайнозое изменений не произошло. Аналогичны данные и по классам растений; здесь также разделяются нижний палеозой, верхний палеозой, мезозой и кайнозой, лишь отмирания классов почти не происходило.
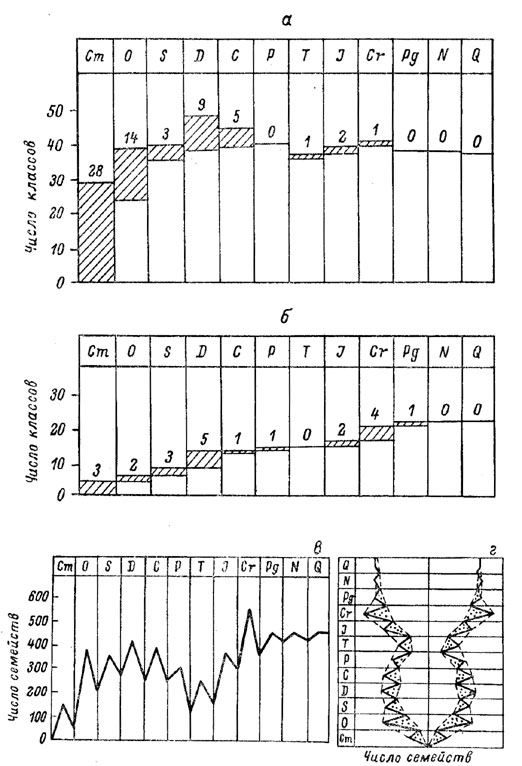
Рис. 40. Эволюция живой природы в фанерозое. а и б - число классов соответственно животных и растений в различные периоды фанерозоя (заштрихованные части колонки и цифры над ними указывают число новых классов, появившихся заданный геологический период); в и г - число семейств беспозвоночных животных.
В качестве дополнительной иллюстрации на рис. 40, в приведен график числа семейств беспозвоночных животных (без хелицеровых и трахейнодышащих членистоногих и некоторых других; всего учтено 1831 семейство). Он демонстрирует стабильный уровень в палеозое, спад в триасе, бурный рост в мезозое, максимум в меловом периоде и затем некоторый спад к стабильному уровню кайнозоя. Таким образом, данные о разнообразии форм организмов как по их крупным группам (рис. 40, а), так и по средним (рис. 39 и 40, в), несомненно, дают основания для разделения фанерозоя на палеозой и мезокайнозой; некоторые основания есть и для разделения мезозоя и кайнозоя и, может быть, для деления палеозоя на нижний и верхний. Однако дальнейшее подразделение на периоды по различным группам организмов пришлось бы осуществлять по-разному, аналогично тому, как периодизация по тектоническому признаку была бы разной в различных регионах.
Обобщение палеонтологических материалов дает сведения и о темпах эволюции форм организмов, т. е. о промежутках времени, потребовавшихся на формирование групп различных уровней. Так, типы животных сформировались за время порядка 1.3 млрд. лет - от среднего протерозоя до кембрия. По оценке выдающегося советского биолога Л. А. Зенкевича, в фанерозое формирование классов занимало время порядка сотен миллионов лет, отрядов - десятков миллионов лет, семейств - порядка десяти миллионов, родов - миллионы, видов - сотни и десятки тысяч лет. По мере развития жизни на Земле темпы эволюции, по-видимому, возрастают, так как организмам надо успевать приспосабливаться ко все усложняющемуся окружению. Так, в плейстоцене сложность условий усугублялась резкой изменчивостью климата и отмечалась быстрая смена старых видов новыми. Известен пример разведения кроликов на островке Порто-Санто в XV столетии, приведшего всего за 400 лет к формированию нового вида мелких кроликов, не дающих потомства при скрещивании с кроликами европейского вида.
Эволюция форм организмов происходит, во-первых, вследствие мутаций (вызываемых физико-химическими воздействиями случайных изменений в нуклеинокислотных носителях наследственности) и, во-вторых, вследствие отбора тех мутантов, которые оказываются способными к адаптации (т. е. к приспособлению) к окружающей их живой и неживой природе. В большинстве случаев способными к адаптации оказываются мутанты с полезными усложнениями организмов; полезные упрощения возникают гораздо реже, и жизнь в целом эволюционирует от простого к сложному. Иногда же возникают особенно удачные формы организмов, столь хорошо приспособленные не только к среде, окружавшей их в период образования, но и к широкому спектру ее изменений, что им удается просуществовать чрезвычайно долго, до сотен миллионов лет включительно. Иначе говоря, высокая мера адаптированности приводит к низкой мере эволюционирования (и наоборот).
Приведем ряд примеров «живых ископаемых» - высокоадаптированных форм организмов, просуществовавших очень большие промежутки времени. Водоросли типа синезеленых, сформировавшиеся еще в начале архея, существуют и сейчас. В подтипе сосудистых растений псилофитов, силуро-девонских первых поселенцев суши, сейчас имеются 3 вида, в подтипе плаунов - 4 рода (один из них - ликоподий); существующие с девона подтипы хвощей и папоротникообразных широко распространены и теперь. От появившегося в карбоне отряда голосемянных растений гинкго сейчас остался один представитель, выращиваемый как декоративное растение в Японии и Китае.
С кембрия известны процветающие и поныне простейшие животные класса саркодовых, подклассов корненожек (отряд фораминифер) и радиолярий. До сих пор распространены почти все классы и отряды кембрийских губок. Из известных с раннего кембрия беззамковых брахионод до наших дней сохранились 3 семейства - лингулиды, кранииды и дисциниды, в том числе известный с девона род лингула. Распространенные ныне роды двустворчатых моллюсков нукула, леда, модиолус, острея, лима и авикула существуют с кембрия. Считалось, что моллюски класса однокрышечковых существовали лишь с кембрия по девон, однако в 1952 г. датское исследовательское судно «Галатея» подняло со дна океана, с глубины 4 км, живых однокрышечковых моллюсков, которых назвали неопилинами; они близки к кембрийскому роду триблидиум. До наших дней сохранились головоногие моллюски мелового рода наутилусов кембрийского подкласса четырехжаберных (наутилоидей). Дожили также иглокожие раннеюрского отряда артикулят силурийского класса морских лилий и 3 рода класса крыложаберных, к которому принадлежали граптолиты кембрия-карбона. Сохранились листоногие ракообразные щитни триасового вида триопсканкриформис (это совершенно уникальный случай двухсотмиллионолетнего существования вида!), кембрийский подкласс ракушечковых ракообразных остракод, членистоногие известного с силура отряда мечехвостов кембрийского подтипа клешнеусых, а также представители известного с карбона отряда стрекоз.
До сих пор в Мировом океане процветают представители появившегося в девоне подкласса пластинчатожаберных рыб - акулы; многие из современных семейств акул существуют с юры.
С раннего девона известны два надотряда лопастеперых рыб - двоякодышащие и кистеперые. От первых сейчас осталось всего 6 видов (пермского отряда рогозубообразных), некоторые из них - довольно крупные рыбы (до 2 м), способные к длительной спячке. Кистеперые отряда рипидистий, породив земноводных, вымерли в карбоне. Другой же их отряд целаканты считался вымершим в конце мела. Однако в 1938 г. 1 экземпляр живого целаканта неожиданно попался в сети южноафриканского траулера, что явилось одной из крупнейших биологических, сенсаций XX в. Этот вид был назван латимерией по фамилии хранительницы местного музея М. Куртенэ-Латимер. В результате героических 14-летних усилий профессора Дж. Л. Б. Смита (см. его книгу [51]) удалось найти постоянное место обитания целакантов - 100-метровые глубины в районе Коморских островов к северу от Мадагаскара; интересно, что «биологическая ниша», в которой скрывались целаканты, была столь ограниченной в пространстве. О процветании вида надо судить, по-видимому, как по количеству особей, так и по их размерам. Целаканты малочисленны (вероятно, при их высокой адаптированное многочисленное потомство ненужно), но это красивые, сильные и крупные рыбы длиной до 2 м и весом до 100 кг (рис. 41). Сейчас их лов производится лишь поштучно по заказам музеев. Один экземпляр был доставлен в СССР экспедицией Института океанологии АН СССР.
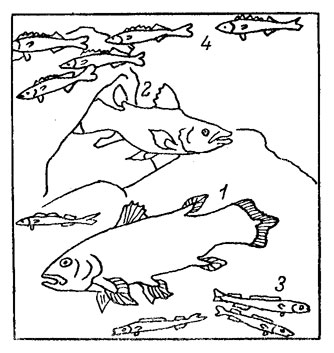
Рис. 41. Латимерия (1, 2) и полуглубоководные рыбы алет (3) и рувета (4).
Из современных амфибий наиболее древними (юрскими) формами являются гладконогиелягушки Северной Америки и Новой Зеландии; из рептилий - черепахи, мало изменившиеся со времен триаса, а также единственный вид, оставшийся от триасового отряда клювоголовых, - гаттерия; из млекопитающих - однопроходные яйцекладущие, которых осталось 5 видов - утконос, два вида австралийских ехидн и два вида новогвинейских проехидн.
Как только что отмечалось, одним из признаков процветания вида можно считать большие размеры особей, тенденции к гигантизму. Они, конечно, свидетельствуют о хорошей адап-тированности вида к окружающей его среде, но такая адаптированность вряд ли устойчива (т. е. гигантизм вряд ли полезен для адаптации к изменениям окружающей среды, поэтому представляется, что гиганты, вообще говоря, не должны быть особенно долговечными).
Самыми крупными организмами на Земле являются бурые водоросли ламинарии, достигающие в длину нескольких сотен метров. Большие размеры имели древовидные плауны, хвощи и папоротники верхнего палеозоя. Из современных деревьев самые крупные - это известные с верхней юры голосемянные секвойи, достигающие в высоту более 110 м и живущие тысячелетиями.
Гиганты встречаются даже среди самых .примитивных животных: у современной сцифоидной медузы цианеи диаметр колокола достигает 4 м, а длина щупалец - 30 м. Мы уже упоминали гигантских головоногих моллюсков наутилусов ордовика с прямыми раковинами 6-метровой длины, меловых аммонитов с закругленными раковинами 3-метрового диаметра, современных кальмаров архитевтисов длиной 15-20 м и более и весом в несколько тонн - это самые крупные из беспозвоночных животных; назовем еще крупнейших из двустворчатых моллюсков - тридаки с полутораметровыми раковинами весом в несколько центнеров.
Выше упоминались крупнейшие из членистоногих - 2-метровые хищные ракоскорпионы нижнего палеозоя гигантостраки. Самые крупные современные рыбы - это планктоноядные китовые акулы длиной до 20 м и более; их хищные родственники - белые акулы кархародоны могут иметь в длину более 11 м, а их плиоценовые предки кархародоны- мегалодоны достигали 30 м; они, по-видимому, были крупнейшими хищниками за всю историю Земли. Самые большие сухопутные хищники - 15-метровые меловые тираннозавры - были вдвое короче.
Абсолютные рекорды веса среди наземных животных принадлежали травоядным динозаврам - 50-тонным брахиозаврам и 30-тонным бронтозаврам.
Самые большие современные птицы - это африканские страусы ростом до 2.7 м и весом до 70-90 кг.
Из млекопитающих гигантизма достигали даже такие примитивные животные, как неполнозубые: неогеновые ленивцы мегатерии были величиной со слона, а броненосцы глиптодонты - с носорога; со слона были также копытные палеоцена и эоцена уинтатерии. Самые крупные наземные животные наших дней - это африканские слоны ростом до 4 м и весом 7.5 т, самые же крупные животные всех времен - это современные синие киты, достигающие в длину более 33 м, а по весу свыше 150 т (сейчас они практически уже истреблены человеком).
Почти все гиганты прошлого вымерли, как и вообще вымерли многие виды, роды, семейства, отряды и даже классы (рис. 40, а). Выше уже отмечалась неравномерность эволюции разнообразия форм организмов: на границе силура и девона исчезло большинство форм граптолитов, трилобитов, цистоидей и наутилоидей; на границе перми и триаса резко сократилась палеозойская флора плаунов, хвощей и семенных папоротников, окончательно вымерли трилобиты, цистоидей, древние морские ежи, многие виды брахиопод, гигантостраки, гониатиты, большинство стегоцефалов и многие другие формы. Наконец, на границе мела и палеогена исчезли аммониты, белемниты, многие брюхоногие моллюски, динозавры, плезиозавры, птеродактили, зубастые птицы. Эти «роковые» границы отмечены на рис. 37 жирными линиями. Причинами такого вымирания были, конечно, резкие изменения окружающей живой и неживой природы (в том числе климата) и, вероятно, в первую очередь такие изменения в живой природе, как появление врагов, опасных для данной группы организмов (например, появление животных, активно поедающих яйца динозавров). По-видимому, нет каких-либо оснований считать, что вымирание тех или иных групп организмов имело характер мгновенных катастроф, подобных лучевым ударам от взрывов сверхновых звезд, - скорее это было постепенное вырождение популяций в течение ряда поколений.
Важное значение для решения вопросов истории Земли могут иметь данные палеобиогеографии о распределении по поверхности Земли и миграциях тех или иных групп организмов в различные периоды времени. Одним из примеров могут служить свидетельства палеоботаники и палеонтологии в пользу единства материков Гондваны в пермо-триасовое время. В южных областях Африки (ЮАР - Трансвааль) и Южной Америки (Аргентина), в Индии и в Антарктиде (Трансантарктические горы) в перми была распространена удивительно однородная растительность глоссоптерид (глоссоптерисы, гангамоптерисы, пара-каламиты и др.), очень резко отличавшихся от флоры континентов северного полушария (частично проникавшей лишь в северные районы Гондваны). В триасе Южной Африки, Южной Америки и Индостана найден ряд сходных лабиринтодонтов и четвероногих рептилий. В 1970 г. американские палеонтологи во главе с Д. Эллиотом обнаружили останки сходных животных в Антарктиде (около ледника Бирдмор) и среди них - останки типичной для Южной Африки и известной также в Индии небольшой, величиной с собаку, коротконогой нижнетриасовой рептилии листрозаурус.
Другим примером аргументации в пользу существования Гондваны, уже упоминавшимся выше, служит расселение млекопитающих в кайнозое. Появившиеся, вероятно, еще в конце триаса однопроходные и зародившиеся в начале мела, по-видимому, на материках северного полушария, сумчатые были оттеснены плацентарными в Южную Америку и Австралию. «Мост» в последнюю вскоре «разрушился», так что из плацентарных проникнуть в Австралию успели лишь мелкие грызуны и летучие мыши; в возникшей изоляции неполнозубые смогли уцелеть, а сумчатые породили почти все разнообразие австралийских млекопитающих.
В эоцене от полуобезьян произошли широконосые обезьяны Америки и узконосые обезьяны Африки и Евразии - «мостов» между этими группами континентов, видимо, тогда не существовало.
Таким образом, к неогену сформировались четыре центра развития млекопитающих:
1) сумчатые и однопроходные в Австралии;
2) сумчатые, неполнозубые и широконосые обезьяны в Южной Америке;
3) разнообразная фауна Евразии и Африки;
4) менее богатая фауна Северной Америки (без мастодонтов, оленей, обезьян, с большим количеством копытных и меньшим количеством хищников). Согласно палеозоогеографическим данным, «мост» между Азией и Северной Америкой образовался в позднем миоцене (и просуществовал до голоцена), а между Северной и Южной Америкой - в среднем плиоцене (существует он и сейчас).
В заключение изложим вкратце историю семейства гоминид, в котором ныне имеется один вид - Homo sapiens (человек разумный). По-видимому, самыми ранними из гоминид были миоценовые рамапитеки. Впервые их останки (фрагменты верхней челюсти кениапитека) найдены в 1961 г. Луисом Лики в Африке в слое возрастом около 14 млн. лет у Форта Тернан, в области Кенийского рифта, на восточном берегу оз. Виктория. Фрагменты челюсти свидетельствовали о ряде признаков человека - укороченном профиле лица, широкой зубной дуге, небольших зубах. Вскоре после этого Э. Симоне отыскал в музеях фрагменты костей рамапитеков, найденные ранее в слоях возрастом 10-12 млн. лет, в южных предгорьях Гималаев - холмах Сивалик (отсюда и название рамапитеки - по имени индийского мифологического героя Рамы). Затем аналогичные останки были найдены в китайской провинции Хунань, в Юрских горах, в Центральной Испании и, наконец, опять в Кенийском рифте в слоях формации Нгороро около оз. Баринго, возраст которых около 10 млн. лет. Все это, вероятно, свидетельствует о широком распространении рамапитеков в миоценовом поясе саванн, тянувшихся от Африки через всю Азию.
Следующим звеном были плиоценовые австралопитеки. Впервые их останки (череп детеныша) найдены в 1924 г. Р. Дартом в образцах камней, собранных рудокопом в дер. Таунг, на краю пустыни Калахари в Южной Африке (отсюда и название «австралопитеки» - южные обезьяны). Позже Р. Брум и Р. Дарт нашли на территории Трансвааля черепа и кости взрослых австралопитеков - существ с обезьяноподобными черепами и лицами, но с похожими па человеческие скуловыми костями, сочленениями челюстей и зубами, с объемом мозга 650 см3, большим, чем у всех известных нам обезьян.
В послевоенное время сенсационные находки костей и каменных орудий австралопитеков (заставившие нас понять, что люди гораздо древнее, чем это считалось ранее) были сделаны все в том же Кенийском рифте, оказавшемся подлинной прародиной человечества. Первые из этих сенсационных находок совершены Луисом Лики в 1959-1960 гг. в Олдувайском ущелье в Танзании (см. его статью [52]). Сначала в слое возрастом около 1.75 млн. лет был найден череп без нижней челюсти австралопитека, названного зинджантропом (Зиндж - древнеарабское название Восточной Африки), а затем в чуть более древнем слое - кости презинджантропа, имевшего объем мозга 650-680 см3, и орудия его труда (грубо оббитые гальки), за которые он был назван Homo hahilis (человек умелый). Позже на западном побережье оз. Рудольф, у холма Лотегем, были найдены наиболее древние останки австралопитеков, возраст которых 5-5.5 млн. лет (период существования австралопитеков тянется от этого времени до 700 тыс. лет тому назад); у Канапои обнаружен обломок плечевой кости австралопитека возрастом около 4 млн лет; в долине р. Омо, в толще формации Шунгуро, найдены многочисленные останки австралопитеков возрастом от 3.5 до 1.3 млн. лет и кварцевые орудия возрастом около 2 млн. лет, а на другом участке этой долины - кости возрастом около 2.75 млн. лет.
В 1968-1972 гг. сын Л. Лики, Ричард, нашел на восточном берегу оз. Рудольф останки не менее 90 особей австралопитеков, а в пункте северовосточная Кооби-Фора в слое возрастом около 2.6 млн. лет - каменные изделия, отщепы из лавы и кварца; это самые древние из известных нам орудий человека. В том же пункте, вислое возрастом около 9 млн лет, найден удивительный «череп 1470» без надглазничного валика и наружного рельефа (которые все еще сохранялись у последующих архантропов и палеоантропов) и емкостью мозга 810 см3, большей, чем у всех австралопитеков. Там же найдены фрагменты черепа ребенка, кости ног и зубы гоминид, близких по типу к обладателю «черепа 1470». Если эти гоминиды - прямые предки современных людей, то являются ли австралопитеки, а может быть, и архантропы и даже палеоантропы боковыми ветвями, тупиками эволюции?
В период с 1 млн. лет до 350 тыс. лет тому назад в Африке и Евразии существовали архантропы (их мозг имел объем около 1000 см3, большое число извилин и развитую долю, управляющую речью) с выдающейся вперед нижней частью лица, без подбородка, с массивными надглазничными валиками; они создавали каменные орудия нижнего палеолита (шелльской культуры - по г. Шелль близ Парижа, и ранней ашельской культуры - по местности Сент-Ашель на севере Франции) и умели пользоваться огнем. К ним относятся: питекантроп (обезьяночеловек) с объемом мозга 940 см3, останки которого были найдены еще в 1891 г. в плейстоцене Восточной Явы; синантропы (т. е. китайские люди) с объемом мозга 850-1300 см3, многочисленные фрагменты скелетов которых возрастом около 500 тыс. лет найдены в начале 20-х годов текущего столетия в пещерах около Пекина; олдувайские питекантропы, останки которых возрастом около 1.1 и 0.5 млн. лет нашел Л. Лики; атлантропы (от, гор Атлас в Африке), останки которых (три нижние челюсти и теменная кость), а также каменные орудия были найдены в 1954-1955 гг. в Алжире; гейдельбержец, нижняя челюсть которого найдена около немецкого города Гейдельберга вместе с костями львов и носорогов; люди миндельского времени со стоянки Вертешсёлёш близ Будапешта, открытой в 1962 г. М. Печем и исследованной в 1963-1967 гг. Л. Вертешем.
350-25 тыс. лет тому назад развивались палеоантропы с мозгом объемом около 1500 см3, как у современных людей, и даже крупнее, хотя и с несколько меньшими лобными долями, с лицом почти без подбородка, толстыми надглазничными валиками и покатым лбом, ростом 150-165 см. Они изготавливали разнообразные каменные орудия среднего палеолита (мустьерской культуры - по пещере Мустье на берегу р. Везер на юге Франции). Во времена палеоантропов начало формироваться человеческое общество. В его основе лежала коллективная охота на крупных животных. Многочисленные останки палеоантропов найдены в Европе, на островах Средиземного моря, в Северной Африке, Передней и Средней Азии. Их типичные представители - неандертальцы, останки которых впервые найдены в пещере долины Неандер около Дюссельдорфа еще в 1856 г.
Около 30-40 тыс. лет тому назад появились неоантропы современного типа - кроманьонцы (по гроту Кро-Маньон во французской провинции Дор-донь, где их останки были еще в 1868 г. найдены Л. Ларте) - с ориньяк-солютрейскими и мадлен-азильскими орудиями верхнего палеолита, а с голоцена - с полированными каменными орудиями эпох неолита, бронзы, железа.
За последние два-три тысячелетия физический облик людей изменился мало (происходит, по-видимому, лишь увеличение их среднего роста, как об этом можно судить, например, по средневековым латам и кольчугам, а в последние десятилетия - и по медицинской статистике; впрочем, все возрастающее число рослых детей - «акселератов» - видно и «невооруженным глазом»). За этот период не изменился, по-видимому, и уровень интеллектуальных способностей людей; во всяком случае, великолепные образцы точности и изящества мышления, например, Аполлония Пергского и ряда других древнегреческих математиков, по нашему мнению, ничем не уступают лучшим современным образцам.
Человеческое общество оказывает все усиливающееся влияние на развитие жизни на Земле, которое началось в форме охоты, рыболовства и истребления опасных для человека животных и вытеснения дикой жизни из населенных районов, а в наши дни идет по пути создания и разведения культурных сортов растений и пород животных и может, в конце концов, привести почти к стопроцентному управлению развитием всех форм жизни. Это может касаться и вида Homo sapiens, развитие которого определяется не столько биологическими, сколько социальными факторами - прежде всего уровнем производительных сил общества и производственными отношениями его членов (и такими надстройками, как складывающаяся стихийно или формируемая сознательно общественная мораль, которая, например, в прошлые столетия вследствие высокой детской смертности поощряла многодетность; в последние же десятилетия при снижении детской смертности, но создаваемых урбанизацией жилищных ограничениях это отношение изменяется).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://nplit.ru/ 'Библиотека юного исследователя'